Kingswood End
Highlights
Kingswood End has yielded a distinctive Visean assemblage of petrifactions and fusain fragments. It differs markedly from the nearby Pettycur assemblage, which is thought to represent a different environmental setting, and is the only known locality for the enigmatic microsporangiate organs Phacelotheca and Melissiotheca. It has considerable potential for future work on the vegetation of this time.
Introduction
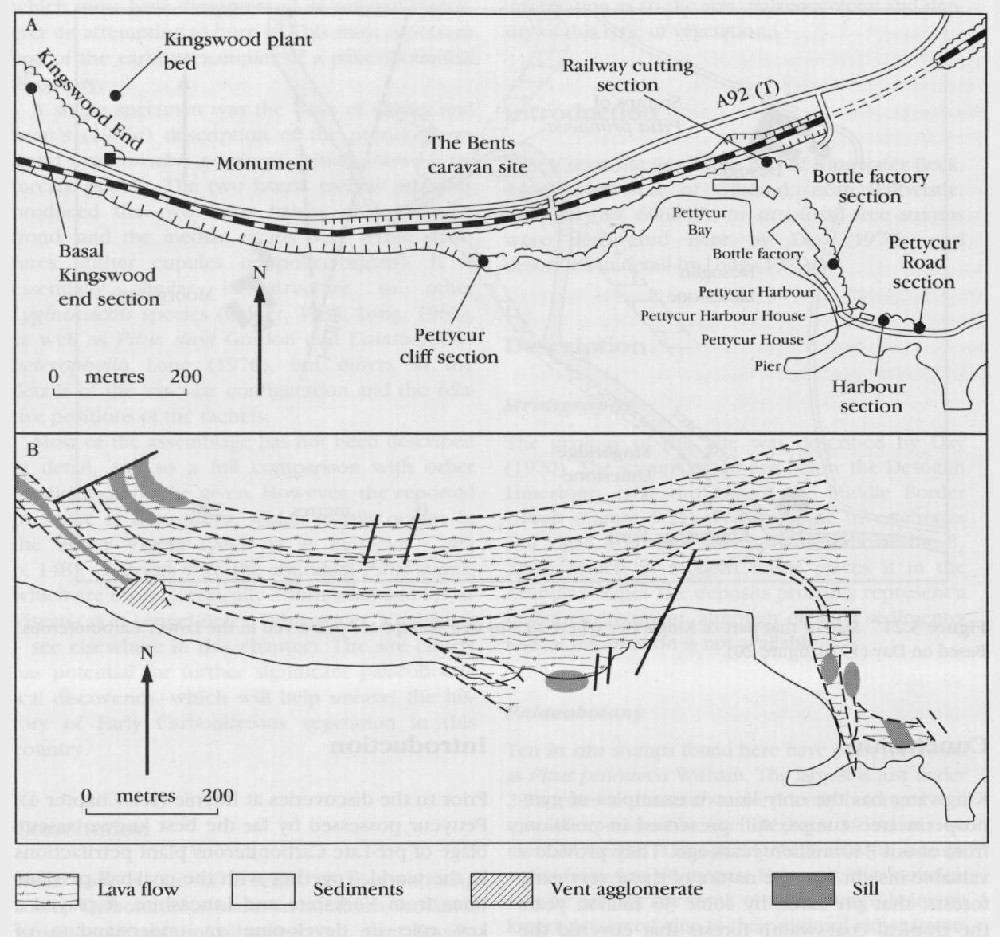
This site lies on the hillside above Pettycur, near Burntisland, Fife
Description
Stratigraphy
The geology of this site is detailed by Scott et al. (1986) and Rex and Scott (1987)
Palaeobotany
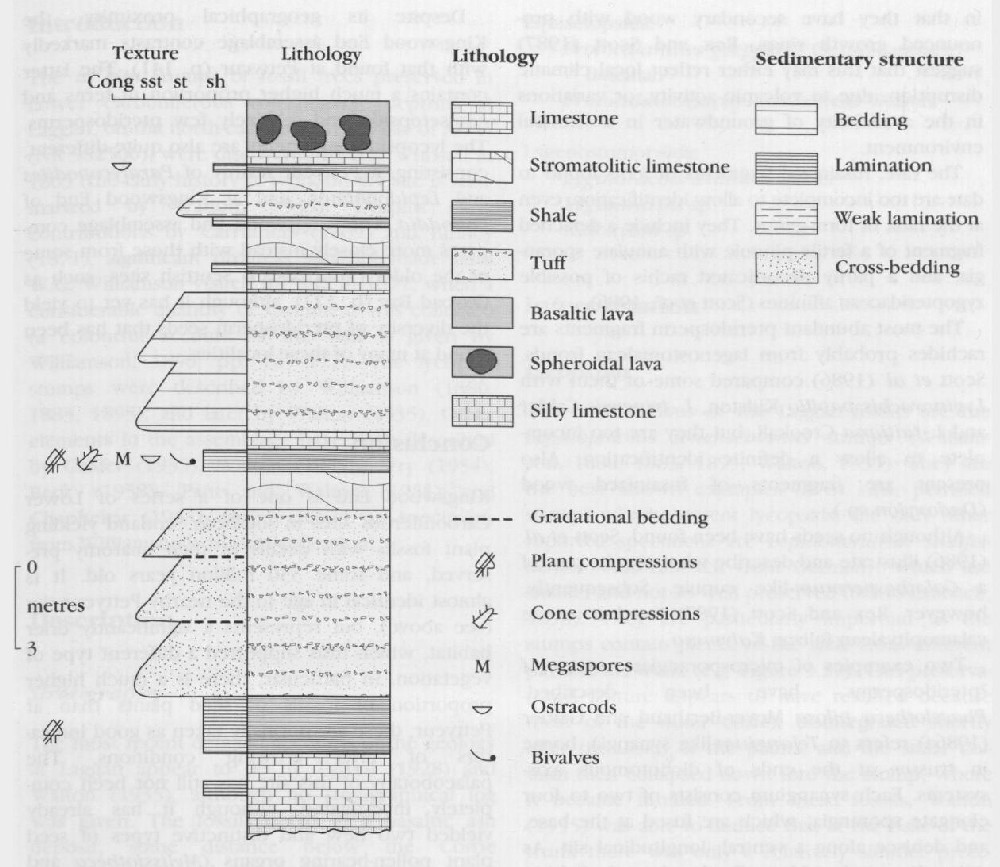
The plant fossils here fall broadly into two groups: petrified lycopsids, and fusainized pteridosperms, ferns and equisetes. Taxa reported to date are as follows:
Lycopsida:
Oxroadia cf. gracilis Alvin
Achlamydocarpon sp.
Equisetopsida:
Protocalamites pettycurensis (Scott) Scott
Filicopsida:
Unidentified fragments
Lagenostomopsida:
Lyginorachis spp.
cf. Calathospermum sp.
Dadoxylon sp.
Uncertain affinities:
Phacelotheca pilosa Meyer-Berthaud and Galtier
Melissiotheca longiana Meyer-Berthaud
Amyelon sp.
In addition, adpressions from a mudstone exposed here have been identified by Rex and Scott (1987) as Lepidostrobus sp. and Lepidostrobophyllum sp.
Interpretation
Oxroadia stems are amongst the most abundant and well preserved fossils from here. They have been interpreted as having lived near the margins of the lake in which the limestone was deposited, and had been subjected to little transportation. Their anatomy compares closely with the well-documented Oxroadia gracilis Alvin from Oxroad Bay (p. 131), except that the larger stems developed a siphonostele, as opposed to a protostele. As pointed out by Scott et al. (1986), however, the Oxroad Bay stems do not reach such a large diameter as the Kingswood End specimens, which may explain the apparently anomalous vascular structure.
Also abundant here are isolated lycopsid megasporophylls. They compare with Achlamydocarpon varies (Baxter) Taylor and Brack-Hanes, but have a rather simpler sporangial wall structure (Scott et al., 1986). Phillips (1979) suggests that Achlamydocarpon may have been borne by giant lycopsids with Lepidodendron-like trunks, but no evidence of the latter has so far been reported from Kingswood End.
The rest of the assemblage consists mainly of fusain fragments. Scott et al. (1986) regard them as the transported remains of plants growing some distance away from the lake, and which had been subject to burning induced by volcanic eruption. They report just two decorticated stems of Protocalamites, which are principally of interest in that they have secondary wood with pronounced growth rings. Rex and Scott (1987) suggest that this may either reflect local climatic disruption, due to volcanic activity, or variations in the availability of groundwater in a stressful environment.
The rare, fusainized fragments of ferns found to date are too incomplete to allow identification, even at the rank of form-genus. They include a detached fragment of a fertile pinnule with annulate sporangia, and a partly decorticated rachis of possible zygopteridacean affinities (Scott et al., 1986).
The most abundant pteridosperm fragments are rachides probably from lagenostomalean fronds. Scott et al. (1986) compared some of them with Lyginorachis papilio Kidston, L. trinervis Calder and L. taitiana Crookall, but they are too incomplete to allow a definite identification. Also present are fragments of fusainized wood (Dadoxylon sp.).
Although no seeds have been found, Scott et al. (1986) illustrate and describe what may be part of a Calathospermum-like cupule. Subsequently, however, Rex and Scott (1987) listed it as the calamopityalean foliage Kalymma.
Two examples of microsporangiate organs of ?pteridosperms have been described. Phacelotheca pilosa Meyer-Berthaud and Galtier (1986a) refers to Telangium-like synangia, borne in trusses at the ends of dichotomous axis-systems. Each synangium consists of two to four elongate sporangia, which are fused at the base, and dehisce along a ventral, longitudinal slit. As pointed out by Meyer-Berthaud and Galtier, Telangium, as currently interpreted, includes a diverse range of microsporangiate organs. They therefore opted to place the Kingswood End specimens in their own, tightly circumscribed form-genus. Virtually all characters point to it belonging to the Lagenostomales, except that the sporangial trusses are rather more complex than in other described species, and the pre-pollen has an ornamented sexine.
A rather more complex structure was described by Meyer-Berthaud (1986) as Melissiotheca longiana. It consists of synangia comprised of 50–150 elongate sporangia, embedded in a lobed, parenchymatous, basal cushion,, which in turn was borne distally on a stalk. The pre-pollen are similar to those contained in Phacelotheca. Meyer-Berthaud pointed out a number of features which again suggest lagenostomalean affinities, but was unwilling to assign it there because of its complexity and the differences in pre-pollen structure.
Despite its geographical proximity, the Kingswood End assemblage contrasts markedly with that found at Pettycur (p. 141). The latter contains a much higher proportion of ferns and equisetopsids, and relatively few pteridosperms. The lycopsid components are also quite different, consisting at Pettycur mainly of Paralycopodites and Lepidophloios, and at Kingswood End of Oxroadia. The Kingswood End assemblage compares more closely instead with those from some of the older (Tournaisian) Scottish sites, such as Oxroad Bay (p. 127), although it has yet to yield the diversity of pteridosperm seeds that has been found at many of these localities.
Conclusion
Kingswood End is one of a series of Lower Carboniferous sites in southern Scotland yielding plant fossils with details of their anatomy preserved, and some 330 million years old. It is almost identical in age to the nearby Pettycur site (see above), but represents a significantly drier habitat, which thus supported a different type of vegetation. In particular, there is a much higher proportion of fossils of seed plants than at Pettycur; these are normally taken as good indicators of drier growing conditions. The palaeobotany of this site has still not been completely investigated, although it has already yielded two new and distinctive types of seed plant pollen-bearing organs (Melissiotheca and Phacelotheca). There is much potential here for new discoveries, which may help in determining aspects of the early evolution of the seed plants.