Pettycur
Highlights
Pettycur is one of the classic Lower Carboniferous palaeobotanical sites and has yielded the most diverse British Visean petrifaction assemblage. It is the type locality for a number of species and is particularly important for lycopsids and ferns; it is also the only known locality for the equisete family Cheirostrobaceae. It is one of the most important palaeobotanical sites in Britain.
Introduction
Prior to the discoveries at Rhynie (see Chapter 4), Pettycur possessed by far the best known assemblage of pre-Late Carboniferous plant petrifactions in the world. Together with the coal-ball petrifactions from Yorkshire and Lancashire, it played a key role in developing an understanding of Palaeozoic plant life. The site
The most detailed investigation of the site was by Gordon (1908b, 1909, 1910a, b, 1911a, b, 1912). Other significant publications were by Benson on the lycopsids, ferns and pteridosperms (Benson, 1908, 1911, 1914, 1922, 1933), as well as some shorter contributions by Kidston (1907, 1908), Bertrand (1907, 1909), Chodat (1912) and Graham (1935).
As at Rhynie, the development of the acetate peel method of investigating petrifactions Joy et al., 1956) had a significant impact on the study of the Pettycur fossils, allowing more detailed reconstructions to be achieved. It proved particularly significant for the ferns, whose complex branching patterns could only be properly resolved by very fine serial sectioning. The most important contributions using the method have been by Surange (1952a, b), Lacey et al. (1957), Chaloner (1958), Holden (1962), Chaphekar (1963) and Chaphekar and Alvin (1972). In subsequent years, there has been little published work on the palaeobotany of Pettycur, although there has been considerable interest in its stratigraphy and sedimentology (Scott et al., 1984; Scott and Rex, 1987; Rex and Scott, 1987).
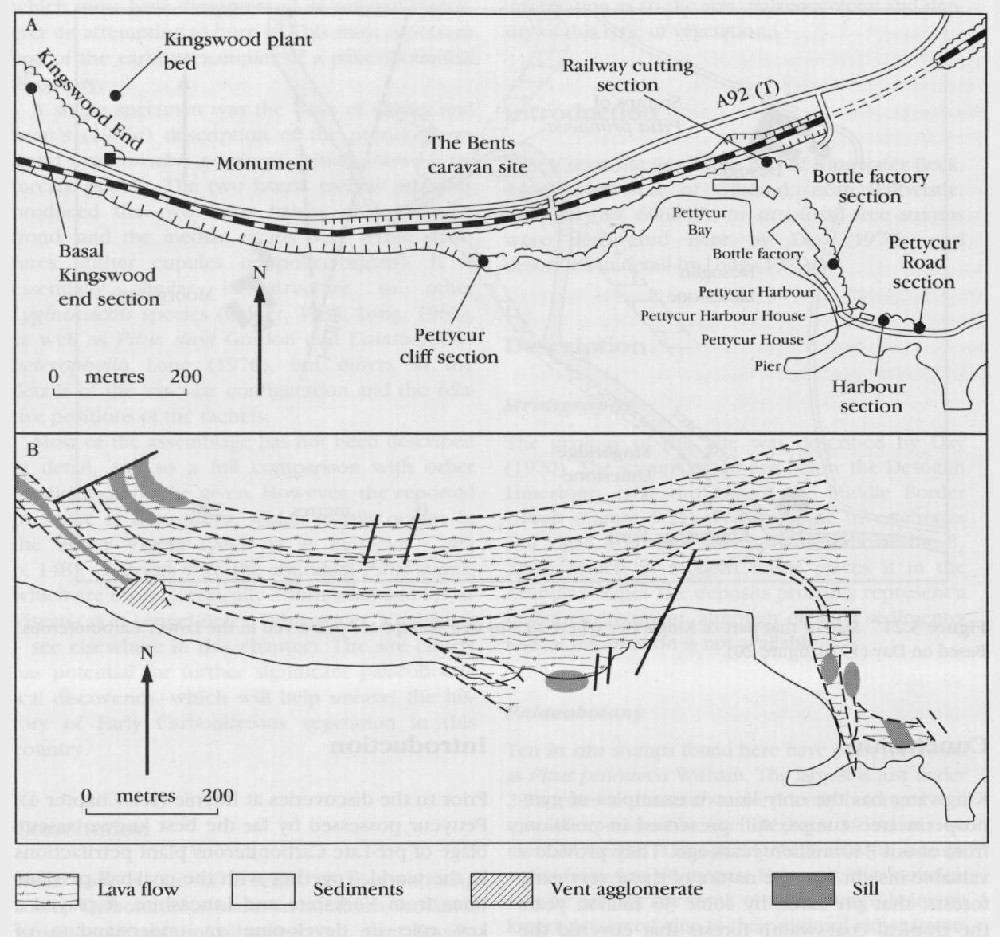
Description
Stratigraphy
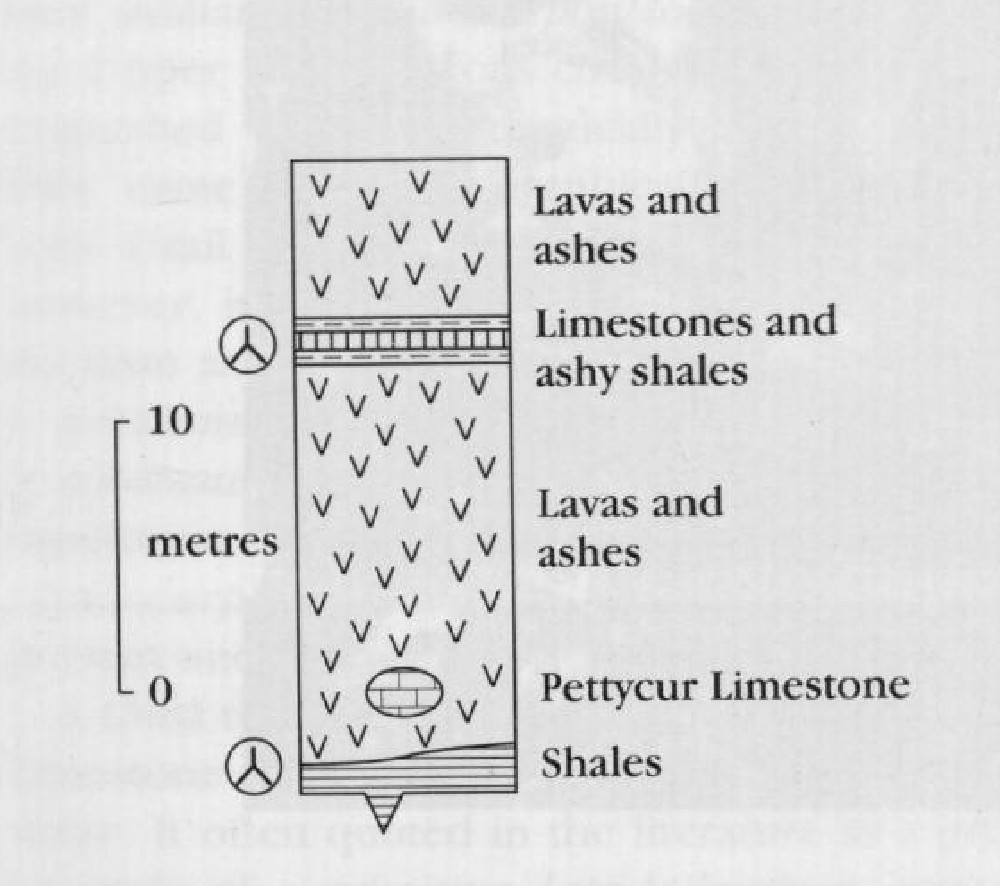
The geology at Pettycur has been extensively discussed by Rex and Scott (1987). The sequence
Palaeobotany
The following form-species preserved as petrifactions and/or fusain have been reported here:
Lycopsida:
Paralycopodites brevifolius (Williamson) DiMichele
Lepidophloios scottii Gordon
'Lepidodendron' pettycurensis Kidston
Flemingites scottii (Jongmans) Brack-Hanes and Thomas
Lepidostrobus cylindricus Gordon M.S.
Lepidocarpon wildianum Scott
Mazocarpon pettycurense Benson
Stigmaria ficoides (Sternberg) Brongniart
Equisetopsida:
Protocalamites pettycurensis (Scott) Scott
Protocalamostachys pettycurensis Chaphekar
Sphenophyllum insigne Williamson
Cheirostrobus pettycurensis Scott
Filicopsida:
Botryopteris antiqua Kidston
Metadineuron ellipticum (Kidston) Galtier
Diplolabis roemeri (Solms-Laubach) Bertrand
Metaclepsydropsis duplex (Williamson) Bertrand
Musatea duplex Chaphekar and Alvin
Stauropteris burntislandica Bertrand
Bensonites fusiformis Scott
Lagenostomopsida:
Heterangium grievii Williamson
Rhetinangium arberi Gordon
Sphaerostoma ovale (Williamson) Benson
Physostoma sp.
Bensoniotheca grievii (Benson) Mickle and Rothwell Amyelon sp.
In addition, Rex and Scott (1987) list as adpressions Lepidodendron sp., Lepidocarpon cf. waltonii Chaloner, Sphenopteris affinis Lindley and Hutton, Sphenopteris sp., Sphenopteridium sp., Adiantites machanekii Stur and Cardiopteridium sp.
Interpretation
Lycopsida
Lycopsid fragments are the most abundant fossils here, particularly in the
Smith (1962c) argued that the stems now called Paralycopodites were intermediate between Lepidodendron and Lepidopbloios, but DiMichele (1980) clearly showed they differed from both these lycopsids in having a smooth-surfaced siphonostele, a homogeneous periderm and cortex, and deciduous branches. He further argued that it could be distinguished by the presence of persistent leaves, although some large Lepidodendron stems have also now been shown to have been leafy (Leary and Thomas, 1989). A closer analogue might be with the adpressions stems known as Ulodendron, as defined by Thomas (1967b). There are differences in the epidermis of the leaves, the stomata being restricted to two bands along each leaf in Paralycopodites (Graham, 1935), whereas in Ulodendron they are more evenly distributed (Thomas, 1967b); but this may just reflect differences at the species level.
Neither DiMichele (1980) nor Thomas and Brack-Hanes (1984) give any opinion as to the taxonomic position of Paralycopodites. However, the likelihood that it bore Flemingites strobili must surely place it at least as a satellite form-genus of the Flemingitaceae, as defined by Thomas and Brack-Hanes.
A second, but less abundant, lycopsid in the swamps produced the stem known as Lepidophloios scottii Gordon (1908b). These stems have a siphonostele surrounding a mixed pith, a feature which is normally regarded as relatively 'primitive'; other Lepidophloios species usually have a fully medullated siphonostele, except in the more distal branches (DiMichele, 1979). Gordon noted that it has leaf cushions similar to those seen in the adpression species Lepidophloios scoticus Kidston, which occurs commonly in the Lower Carboniferous of Scotland (see also Galtier and Scott, 1986b). Sections through stomata on the leaf cushions have been described by Thomas (1974).
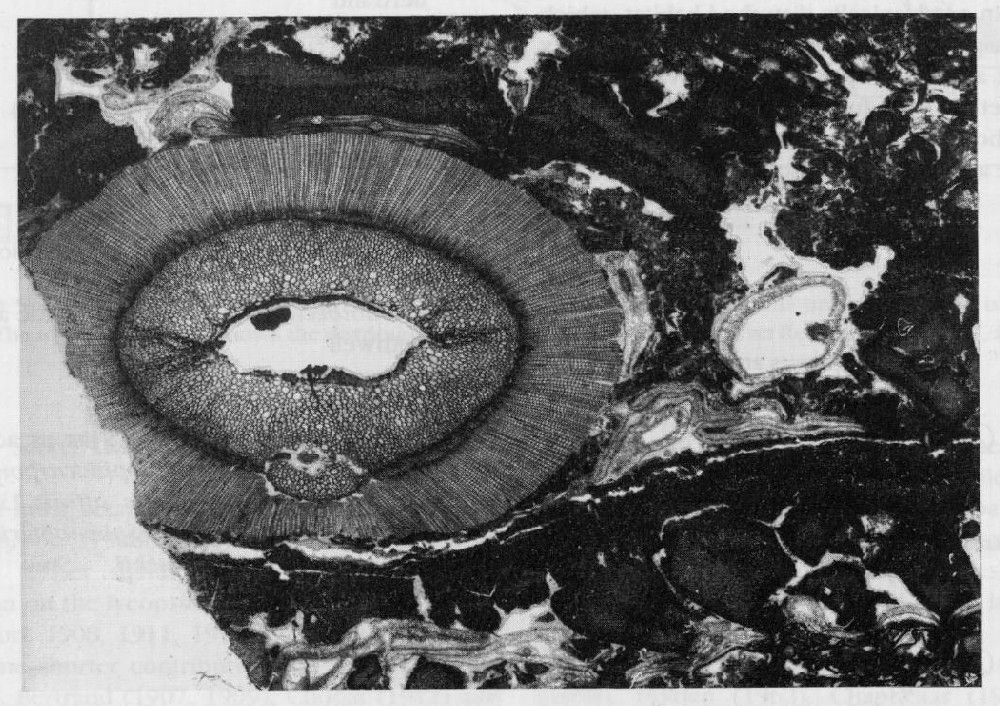
Lepidophloios is thought to have borne mega-sporangiate strobili known as Lepidocarpon (Phillips, 1979; Thomas, 1981b). It is not surprising, therefore, that isolated sporophylls known as Lepidocarpon wildianum Scott (1900, 1901) are associated with Lepidophloios scottii at Pettycur. Being found as isolated sporophylls supports Phillips' (1979) view that they operated as seedlike disseminules. The Pettycur sporophylls are very similar to Lepidocarpon lomaxii Scott from the Upper Carboniferous coal-balls, and Scott distinguished them taxonomically mainly because they came from stratigraphically different horizons. Until complete strobili are found at Pettycur, however, it will be impossible to make any final decision as to the relative taxonomic positions of L. wildianum and L. lomaxii. The relationship of L wildianum to the adpression from the Pettycur mudstones, described by Rex and Scott (1987) as Lepidocarpon cf. waltonii Chaloner, is also at present unclear.
A third type of lycopsid stem from the
The Mazocarpon described by Benson (1908) is only known from incomplete sporophylls, but appears to represent a reproductive strategy similar to that adopted by Lepidocarpon (see above). If correctly assigned to the Sigillariostrobaceae, these sporophylls are the oldest evidence of this family in the fossil record.
A fourth type of lycopsid fructification is Lepidostrobus cylindrica Gordon M.S. Although mentioned in species lists for the site (Gordon, 1914; Walton and Long, 1964), it has never been effectively published and so for the time being must remain a nomen nudum.
Equisetopsida
The remains of two equisetopsid orders have been found at Pettycur: the Equisetales ('archaeo-calamitids') and Bowmanitales ('sphenophylls'). Equisetalean stems are relatively abundant, although Rex and Scott (1987) state that they are often preserved 'in the immature form'. They were originally described as Calamites pettycurensis Scott (1902) and then Protocalamites pettycurensis D.H. Scott (1908–1909). Chaphekar (1963) argued that the differences from 'Archaeocalamites' goeppertii (Protocalamites goeppertii (Solms-Laubach) Bateman) merely reflected different positions within the plant. However, Bateman (1991) has found that the Pettycur stems have consistently fewer primary vascular strands than typical P. goeppertii, such as found at Laggan and Loch Humphrey Burn (Walton, 1949b; Chaphekar, 1963), and that they probably represent smaller, shrubby plants.
Associated with these stems are strobili, Protocalamostachys pettycurensis Chaphekar (1963). Although never found in organic attachment, the structure of the pedicle is very similar to that of the smaller branches of Protocalamites pettycurensis. It differs from Protocalamostachys arranensis Walton found at Laggan (p. 155) in being smaller, having more sporangiophores in each whorl, fewer vascular strands in the pedicle, and containing smaller spores (Bateman, 1991).
The Bowmanitales are rare in the
The order is generally rare in the Lower Carboniferous, although there is evidence of its range extending down to the Upper Devonian (Remy and Spassov, 1959). Petrifactions have also been reported from Saalfeld (Solms-Laubach, 1896) and the Montagne Noire (Galtier, 1970), but the Pettycur specimens remain the best documented Lower Carboniferous examples to date.
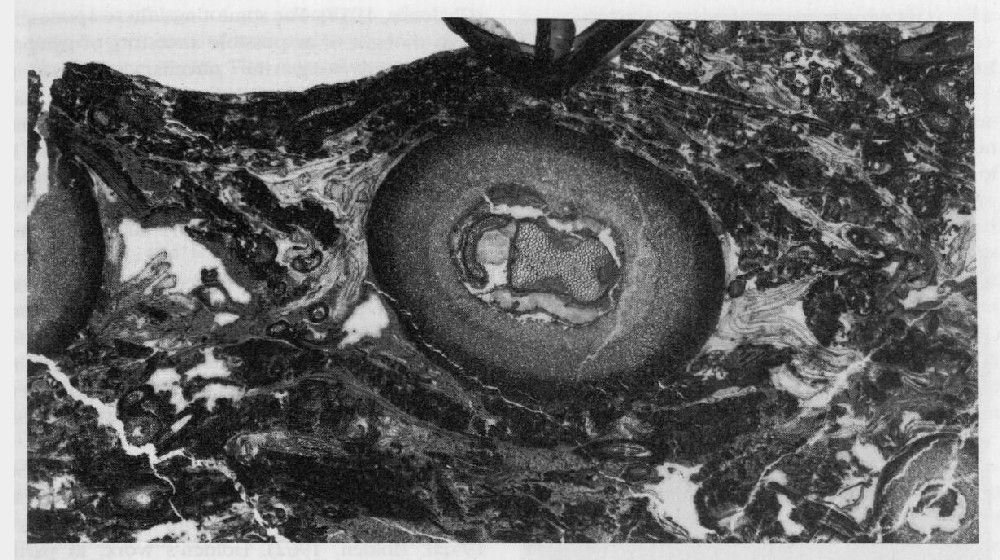
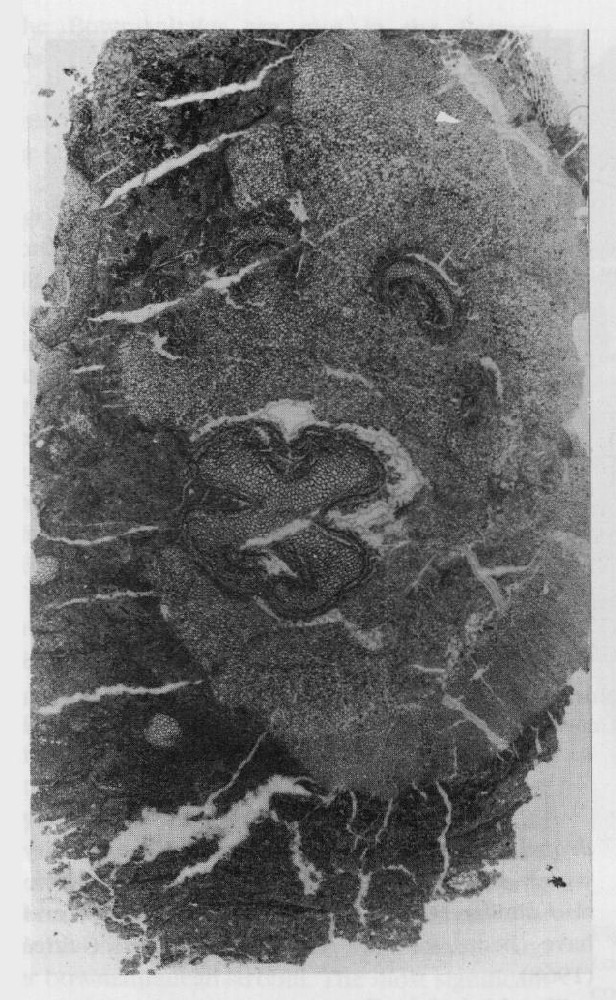
Cheirostrobus pettycurensis
Filicopsida
The ferns fall broadly into two groups: (1) species of Zygopteridaceae (Coenopteridales), which occur mainly in the Zygopterid Limestone; and (2) species of Stauropteridaceae (Coenopteridales) and Botryopteridaceae (Botryopteridales), which occur mainly in the
Zygopterid community
Pettycur is probably the best known British locality for zygopteridacean ferns, and it has provided valuable information on the evolution of the vascular systems of the stem (Gordon, 1911a, b, 1912; Dennis, 1974) and the phyllophores (Gordon, 1911b; Galtier, 1964). They appear not to have been growing in the main peat-swamp, but were probably a pioneer community which would have invaded the areas nearer the volcanic centres during periods of eruptive quiescence (Rex and Scott, 1987). The best understood of the Pettycur species is Metaclepsydropsis duplex (
Of similar habit and anatomy was Diplolabis roemeri
The third of the Pettycur zygopteridacean axes, Metadineuron ellipticum, is only known from fragments of phyllophore (Kidston, 1908; Galtier, 1964, 1970). It is presumed to have been of similar habit to the other members of the family from Pettycur.
Filicopsids of the swamp community
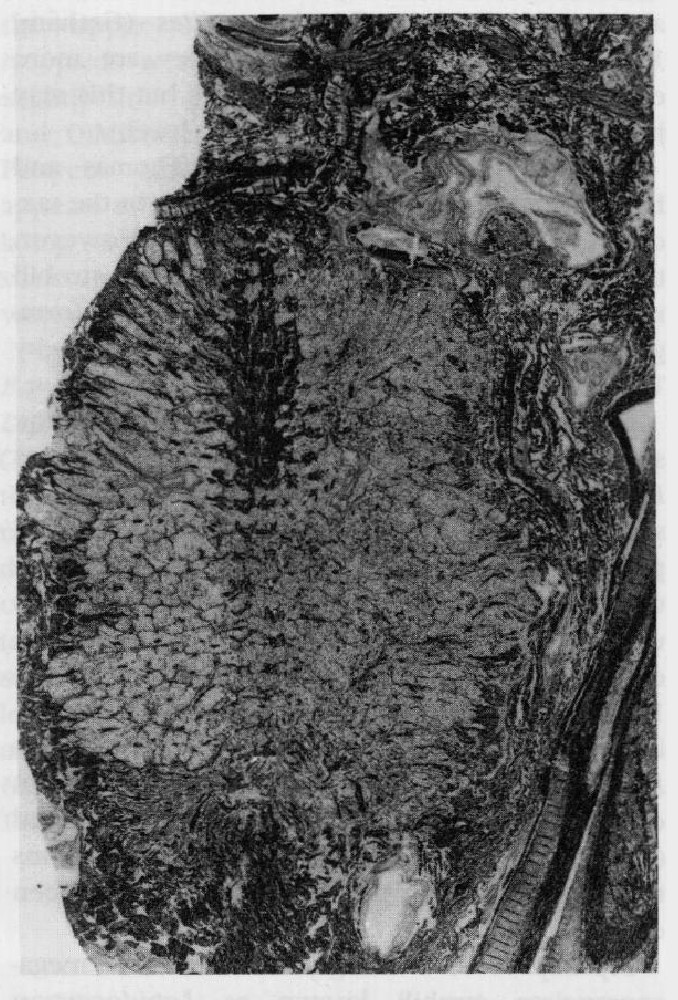
Within the peat-forming swamp, the Coenopteridales were represented by Stauropteris burntislandica (Williamson, 1874b; Bertrand, 1907, 1909). It has also been reported from Roannais in central France (Galtier, 1971), but most work on the species has been based on Pettycur material. Although the overall form of the plant has not been confirmed, it is assumed to have been herbaceous. The fronds were reconstructed by Surange (1952a), and are generally similar to those of the Zygopteridaceae, including the presence of basal aphlebiae (Lacey et al., 1957), but have a less-planated, more primitive aspect.
Of particular interest is the fact that S. burntislandica was heterosporous. The megasporangia had been initially identified as Bensonites fusiformis R. Scott (1908) but the connection with Stauropteris was confirmed by Chodat (1912; see also Surange, 1952a). They are spindle-shaped structures, consisting mostly of parenchyma with a longitudinal vascular strand, but with a distal cavity that opens out to the sporangium apex via a narrow tube. The cavity contains a tetrad of two operative and two aborted megaspores, corresponding to Didymosporites scottii Chaloner (1958) (see also Hemsley, 1990b). Similar dispersed megaspores are known from various Lower Carboniferous localities in Britain (Chaloner, 1958). For some time, these sporangia were thought of as possible ancestors of gymnosperm seeds (e.g. the nucellar modification concept of Andrews, 1961), but this view has now lost favour (see discussion on the Whiteadder earlier in this chapter). Instead, it would appear that it represents a seed-like reproductive strategy, comparable to that achieved by the lycopsids (Lepidocarpon) and equisetes (Calamocarpon).
Microsporangia have not been found in organic connection with S. burntislandica, but R. Scott (1908) reported that structures very similar to Stauropteris oldhamia Binney sporangia occur in close association. Unlike most fern sporangia, they lacked an annulus, and spore-release was achieved via a small stomium.
The Botryopteridales are represented by Botryopteris antiqua, for which this is the type locality (Kidston, 1908; Benson, 1911; Surange, 1952b; Holden, 1962). Holden's work, in particular, has clarified the overall habit of the plant, which seems to have had a creeping stem, giving off erect fronds with digitate, three-dimensional pinnules, and a similarity to the adpression form-genus Rhodeopteridium. A distinctive character of the fronds is the frequent presence of small plantlets, which were presumably a means of vegetative propagation (see also Galtier, 1969), a feature generally rare in the Filicopsida. The sporangia were borne laterally to the ultimate segments of the frond (Galtier, 1967, 1981). They have a transverse annulus and contain spores of the form-genus Granulatisporites.
There has been some disagreement as to the taxonomic position of B. antiqua. It differs from typical Botryopteris (e.g. B. forensis Renault, B. globosa Darrah), which has fertile organs consisting of large clusters of several hundred sporangia. Good (1981) argued that it should therefore be transferred to Psalixochlaena, but this ignores the differences in vascular structure and position of attachment of the sporangia (Holden, 1960; Holmes, 1977, 1981). It is arguable that B. antiqua should be transferred to a different form-genus but, in the absence of a detailed taxonomic analysis of the problem, the traditionally accepted combination is used here.
Lagenostomopsida
Unlike most other Lower Carboniferous petrifaction sites in Britain, Pettycur does not have a diverse assemblage of pteridosperms; there are just two types each of stem and seed, and one of pollen-bearing organ. The most abundant fossils are the stems Heterangium grievii, for which Pettycur is the type and best locality (Williamson, 1873; Williamson and Scott, 1895; Benson, 1933). This is the best known of the 'primitive' group of Heterangium species, which are assigned to the subgenus Heterangium (synonym Euheterangium auct.), and is characterized by a mesarch protostelic vascular system and transverse sclerotic plates in the cortex. The latter feature imparts a transverse bar-pattern to the outer surface of the stems, allowing a correlation with Sphenopteris elegans Brongniart and S. affinis Lindley and Hutton types of foliage preserved as adpressions. These fronds have deeply dissected pinnules, in contrast to the Eusphenopteris fronds borne by Heterangium subgenus Polyangium from the Upper Carboniferous, which have rounded pinnules (Shadle and Stidd, 1975).
The present distribution of H. grievii suggests that it came from a plant that was abundant in the
Associated with H grievii are small Sphaerostoma ovate seeds (Williamson, 1877; Benson, 1914). Although never found in organic connection, Benson argued strongly that they were parts of the same plant. Such seeds have a number of 'primitive' features, such as the integument and nucellus being only partially fused and an abscission layer between the integument and the cupule. However, the salpinx is considerably reduced and there is a well-developed micropyle. It is thus apparently intermediate between the primitive lagenostomalean seeds from the Tournaisian (e.g. Stamnostoma from the Whiteadder — see above) and Lagenostoma from the Upper Carboniferous.
Also in association with H. grievii are some poorly preserved pollen-bearing organs identified by Benson (1922) as Heterotheca grievii (later renamed Bensoniotheca grievii by Mickle and Rothwell (1979)). Consisting of an ovoid cluster of completely fused sporangia, they contrast markedly with the loose clusters of free sporangia associated with other Lower Carboniferous Heterangium stems, and known as Telangium or Telangiopsis (e.g. Jennings, 1976). It is possible, however, that they are merely immature forms of Telangium.
The second type of pteridosperm stem found at Pettycur is Rhetinangium arberi Gordon (1912), for which this is again the type locality. Like Heterangium, these stems have a single protostele, but it is exarch and there is no evidence of sclerotic plates in the cortex.
No seeds have been found in organic connection with Rhetinangium at Pettycur, but Gordon (1910a) reported a poorly preserved example of what seemed to be Physostoma in close association. Gordon's description is only brief and was not accompanied by an illustration. It is perhaps worth noting that in the Tournaisian assemblage at the Whiteadder, there is evidence that Rhetinangium bore Genomosperma seeds.
Walton and Long (1964) reported Amyelon from Pettycur, which may represent pteridosperm roots. However, they have not been described in the literature.
Rex and Scott (1987) list and illustrate a number of pteridosperm foliage fragments preserved as adpressions. However, there is no published taxonomic account of these fossils, and they are thus difficult to judge.
General remarks
This site has yielded the most diverse Visean plant petrifactions known from Britain, and is of great historical significance, having been central to the work of such leading palaeobotanists as W.C. Williamson, D.H. Scott and W.T. Gordon. One of the plant-bearing horizons (the
Conclusion
Pettycur is one of the classic Palaeozoic palaeobotany sites in Britain, yielding anatomically-preserved plant remains about 330 million years old. Together with the peat-floras of the Devonian Rhynie Chert (see Chapter 4) and the Upper Carboniferous coal-balls (see Chapter 6), it was for many years regarded as the best guide to the vegetation of the Palaeozoic. Although many other Lower Carboniferous sites, in this country, France and Germany, are now known to yield anatomically-preserved plant fossils, Pettycur remains of great palaeobotanical significance, particularly for the study of the early fern-like plants and club-mosses. It is also the only known locality for the horsetail family, the Cheirostrobaceae. Despite being one of the oldest known types of equisete, it has by far the most complex reproductive organs ('cones') that have been found in that group of plants, either living or in the fossil record.