Achanarras Quarry
Highlights
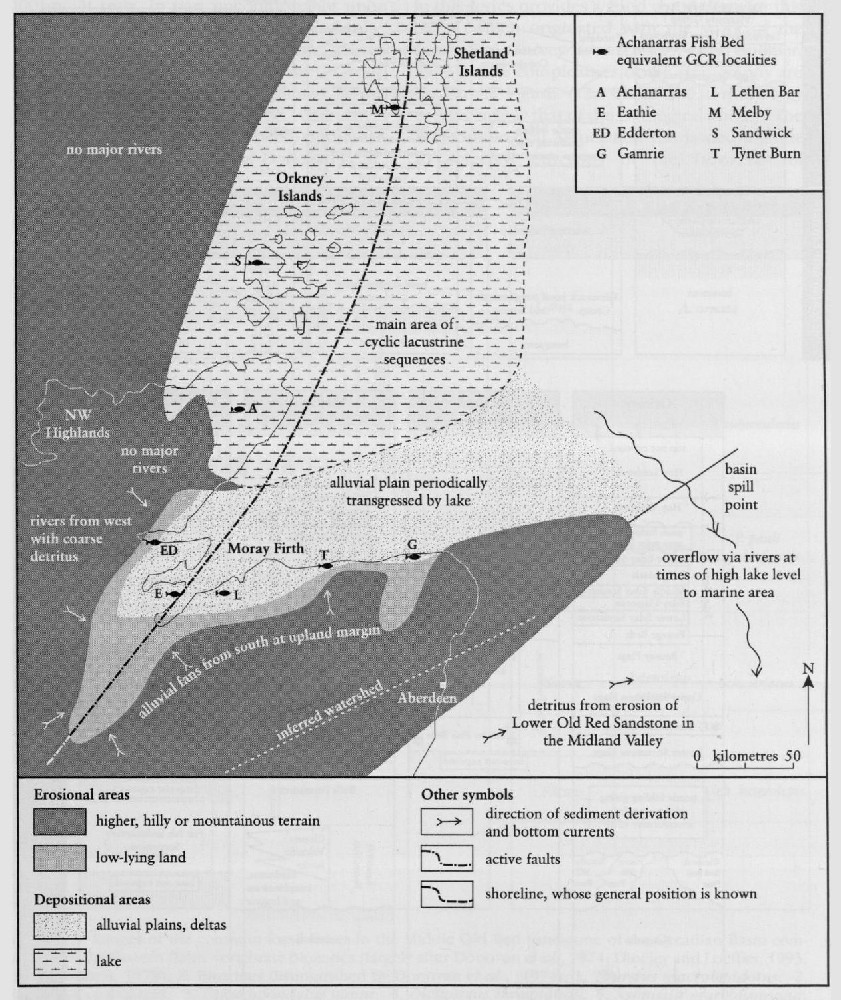
Achanarras in Caithness (Highland) is one of the most famous fossil fish sites in Britain. There are many fish-bearing beds in the quarry, and 15 species have been found in huge abundance there. Achanarras Quarry represents a sample of the rich fish fauna from the south-western quarter of the great Orcadian Lake.
Introduction
The quarry was opened in 1870 (Traquair, 1894a) and since then has yielded many fossil fishes, which may be seen in museums not only throughout Britain, but also elsewhere in the world. By 1914, very little quarrying had taken place for several years (Crampton and Carruthers, 1914). Since that time it has yielded fossils to several collectors, notably C. Forster-Cooper (1937), and has been quarried recently for scientific studies (Trewin, 1986). A field excursion guide has recently been published (Trewin and Hurst, 1993). The quarry is now flooded and the fish bed crops out under water
Description
The Achanarras Limestone is a relatively pure limestone laminite at the top of the
The fish bed at Achanarras is about 1.95 m thick, and can be examined in the north-cast corner of the quarry. The quarry was drained in 1980 and a representative continuous column of rock collected through the full thickness of the fish bed. Etched laminae from the edges of fish-bearing slabs from museum collections could be matched with the patterns exhibited by the total section, and enabled Trewin (1986) to produce detailed information about changing species abundances in the fish bed (Trewin, 1986, figs 2, 3). Trewin (1986) found six clearly defined horizons, with marked changes in diversity and relative abundance between them, and the divisions between the faunal horizons frequently coincide with episodes of low fish abundance. Dipterus is abundant at the top and bottom of the bed (Ahlberg and Trewin, 1995); Osteolepis, Glyptolepis, Coccosteus and Cheirolepis appear after Dipterus, Pterichthyodes, Mesacanthus and Palaeospondylus in the second division, but Rhamphodopsis and the larger acanthodians do not enter until near the top of division three.
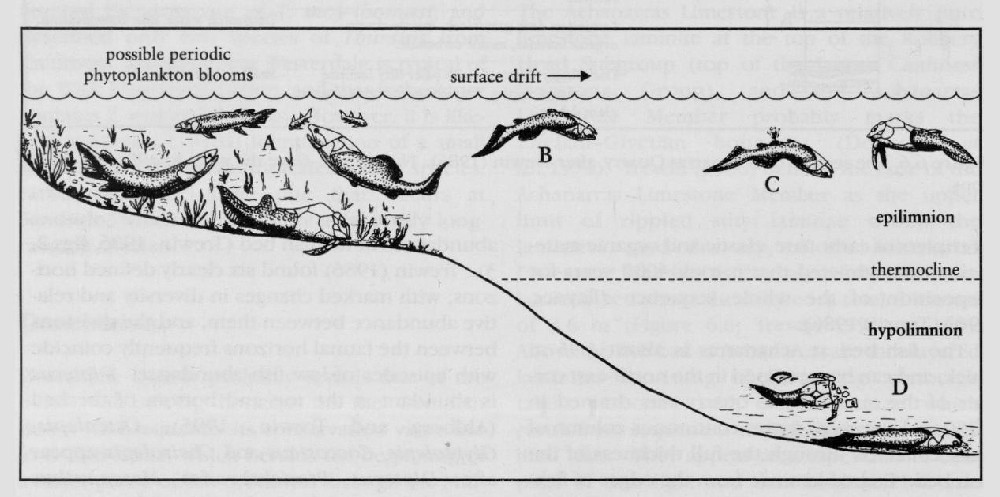
The fossil fishes tend to lie together on certain bedding planes, often with limited signs of disturbance. Many acanthodians, for example, show no signs of putrefaction before burial. Other fossils show evidence of decomposition, as in poorly preserved Dipterus which are bent backwards with bloated bellies, features typical of many dead fish today. Some larger fishes apparently floated around in the upper waters of the lake, buoyed up by decomposition gases in the body chamber, for long enough for parts of them to drop off. Osteolepid specimens are often broken up, and partly scattered around the area of the main body of the fossil
Fauna
Acanthodii: Climatiformes: Diplacanthidae
Diplacanthus striatus
D. tenuistriatus Traquair, 1894
D. (Rhadinacanthus) longispinus (Agassiz, 1844)
Acanthodii: Acanthodiformes: Acanthodidae
Mesacanthus peachi Egerton, 1861
Cheiracanthus murchisoni Agassiz, 1835
Placodermi: Antiarchi: Pterichthyodidae
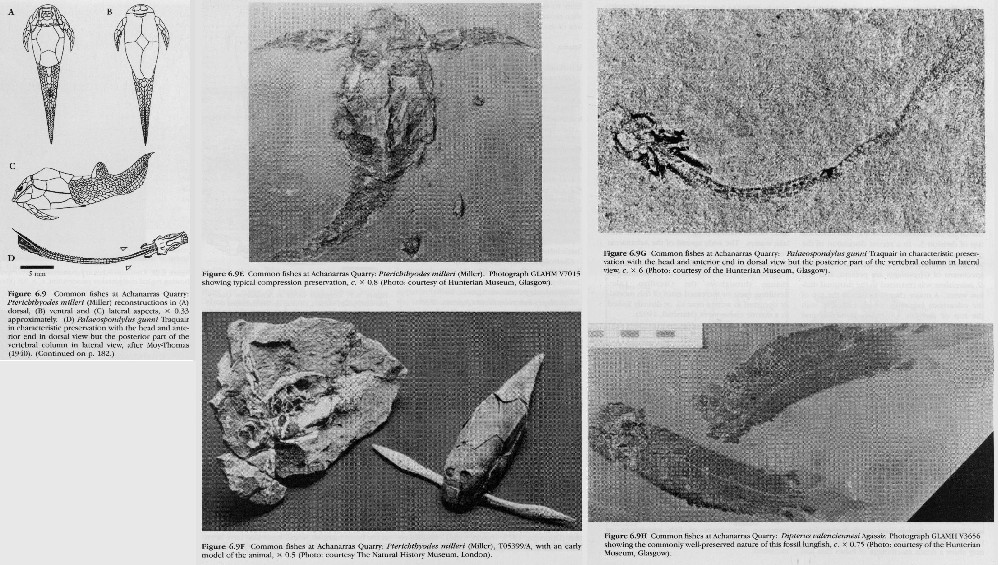
Pterichthyodes milleri Miller, 1841
Placodermi: Arthrodira: Coccosteidae
Coccosteus cuspidatus Miller, 1841
Placodermi: Arthrodira: Homosteidae
Homosteus milleri Traquair, 1888
Placodermi: Ptychtodontida: Ptychtodontidae
Rhamphodopsis threiplandi Watson, 1938
Incertae sedis
Palaeospondylus gunni Traquair, 1890
Osteichthyes: Actinopterygii: Cheirolepidae
Cheirolepis trailli Agassiz, 1835
Osteichthyes: Sarcopterygii:
Osteolepidiformes: Osteolepididae
Glyptolepis paucidens Agassiz, 1844
Osteolepis macrolepidotus Agassiz, 1835
Osteichthyes: Sarcopterygii:
Porolepiformes (Holoptychiida): Holoptyiidae
'Holoptychius' sp.
Osteichthyes: Sarcopterygii: Dipnoi: Dipterida: Dipteridae
Dipterus valenciennesi Sedgwick and Murchison, 1828
These fishes are described in the introduction to the chapter. Achanarras Quarry is the type locality of two species, Rhamphodopsis threiplandi Watson, 1938 and Palaeospondylus gunni Traquair, 1890
Interpretation
Carbonate laminae may have formed as a result of increased pH following photosynthesis, hence causing carbonate (dolomite or calcite) deposition (Donovan, 1975; Trewin, 1986). Studies at Lake Balaton in Hungary indicate that calcite is precipitated at times of high lake level and low evaporation. The laminites from Achanarras, however, are thought to have been caused by a seasonal climate, with each dark organic-rich band representing the silts deposited during the rainy season, and the light band the carbonates that came out of solution during a dry summer. Each light and dark pair can be counted. They have been interpreted as the products of annual algal blooms. Trewin (1986) concluded that the 'calcitic' laminites of the central part of the fish bed represent the period of highest lake level, with greatest faunal diversity because of the stable environments, and the possibility of maximum migration around the lake. He suggested that they are the equivalent of the nodule beds at the basin margins, which must also have been deposited at times of maximum lake extension.
The fossil fishes tend to lie together on certain bedding planes, representing catastrophes that killed off schools of fish. Postulated causes of mass mortality include not only salinity crises, but also storms stirring up toxic deeper waters, algal blooms leading to eutrophication of surface waters, or shock mixing of cold deep waters with the warmer surface water
There is a series of phases of faunal introduction. Dipterus is abundant at the top and the bottom of the fish bed, suggesting that it was the first fish to colonize an area and the last survivor. Dipterus may have tolerated lower oxygen levels than the other fishes (Trcwin, 1986; Ahlbcrg and Trewin, 1995) and was environmentally more hardy than the other fishes. Pterichthyodes, Mesacanthus and Palaeospondylus then appear and become abundant within the calcareous laminites of divisions 2, 3 and 4. Osteolepis, Glyptolepis, Coccosteus, and Cheirolepis all appear in division 2, but Rhamphodopsis and the larger acanthodians do not appear until the top of division 3. In a recent discussion of the palaeoecology of the porolepiform fishes of Scotland, Ahlberg (1992) noted that only adult specimens of Glyptolepis are present, and that G. paucidens was restricted to the marginal shallow water. A major change occurs at the top of the calcareous laminites, which coincides with the top of division 4. Palaeospondylus and Cheirolepis disappear, and there is a major expansion of Coccosteus in the overlying dolomitic laminites. Other species linger on in small numbers. Thus, changes in conditions which gave rise to dolomite, rather than calcite precipitation, were not equally tolerated by all species. Finally, division 6 contains only Dipterus
The Achanarras horizon fauna consists of the same species throughout the area, but significant differences occur in the proportions present at each site, and these may correspond to different ecological areas of the lake. Achanarras Quarry has yielded the greatest number of species, possibly because it was more centrally placed within the lake, whereas sites in the Moray Firth area were on the margin, and contain a sparser fauna
Conclusion
The Achanarras Quarry is flooded for most of the time, so the fish bed may not be exposed above the water. Nevertheless, it is world famous as perhaps the richest Old Red Sandstone fish site in Britain. It represents the richest sampling of Middle Devonian fish diversity in the Orcadian Lake, having been located in quiet open waters, removed from the littoral. Many thousands of specimens of fossil fishes, collected over the years, include well-preserved representatives of at least 15 species. Recent sedimentological work has enhanced the conservation value of the site, in linking high-resolution biostratigraphy with models of ancient lake development, and the fluctuations of the fish populations. Exploratory excavations have proved the immense abundance of fish specimens at Achanarras Quarry, and its potential seems undiminished.