Chapter 2 Fossils of the Chalk and the ecology of the Upper Cretaceous Chalk seas
Fossils of the Chalk
Upper Cretaceous Chalk is remarkable for its diversity of fossils. The ultra-fine fraction (below ten micron size) contains about 30 000 coccoliths (
The superbly preserved fossils seen in museum collections were obtained when quarries were worked by hand. Some of the finest museum specimens (e.g. starfish), are actually cleverly constructed fakes, put together using components from more than one species.
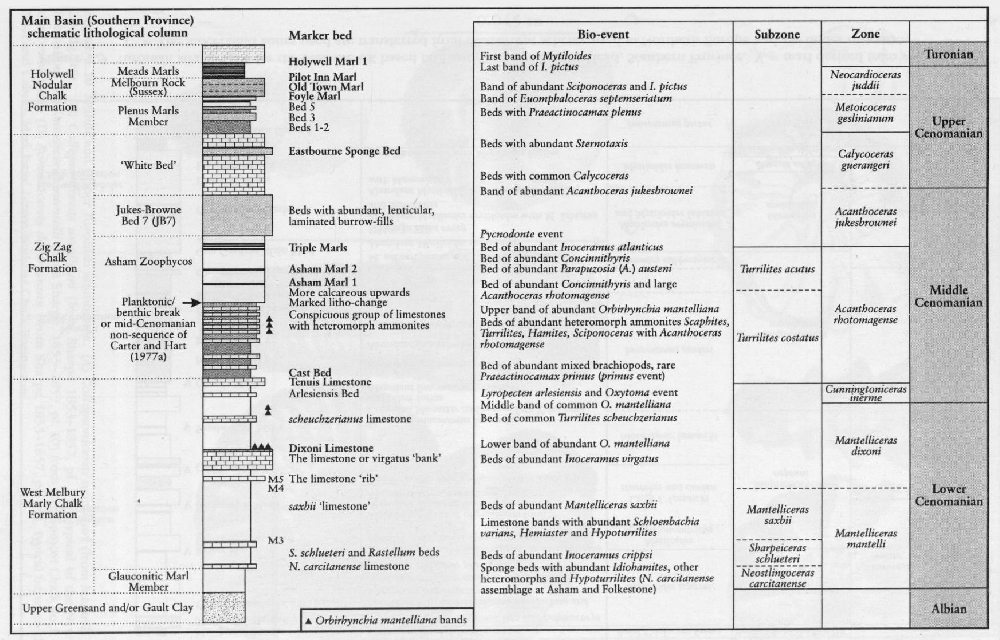
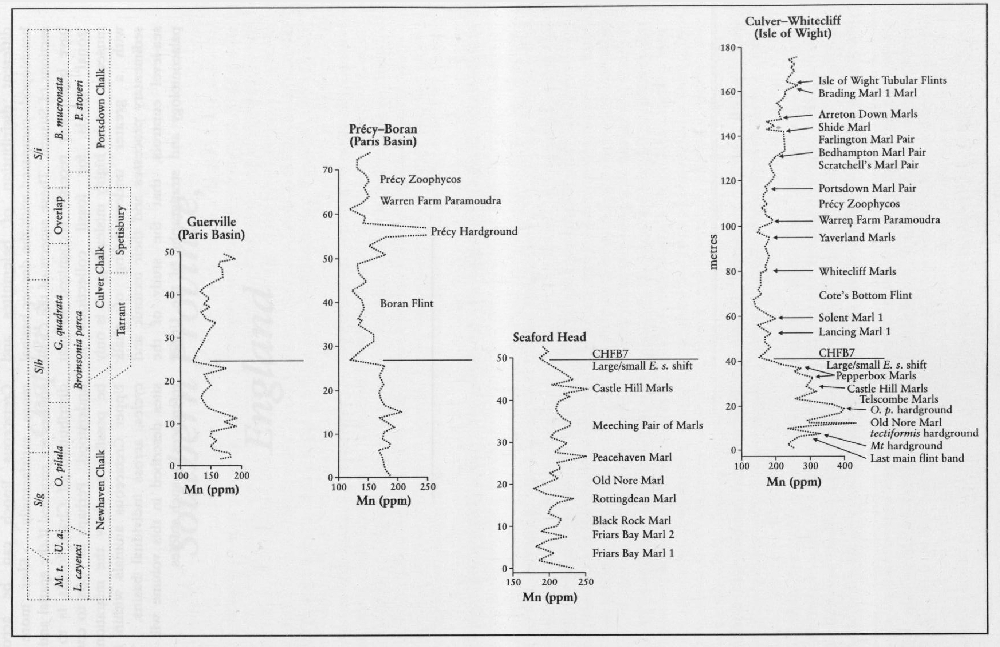
A feature of the Chalk is the cyclic occurrence of more and less calcareous horizons (couplets; see
There are several anomalies in the distribution of fossils in the Chalk. Some horizons are rich in macrofossils while other horizons appear to be almost barren of any diagnostic forms. Diversity of fossils also changes stratigraphically. The higher parts of the Lower Turonian
Preservation of fossils is also variable. Basin margins where shallow-water calcareous sands developed in the early part of the Cenomanian, such as south-east Devon, are areas where originally aragonitic-shelled fossils, particularly ammonites, are better preserved and more abundant than in the regions where deeper-water chalks formed. Similarly, otherwise extremely rare ammonites are found in the Upper Turonian Chalk Rock hardgrounds of the Chiltern Hills (on the Anglo-Brabant Massif). Right across Europe there is a preservational event at the equivalent level of the Chalk Rock. This type of event is found again in the Catton Sponge Bed and the Trimingham Sponge Bed in Norfolk and Northern Ireland. Not all of the sponge beds and hardgrounds in the Chalk show this type of preservation and it is puzzling why some do whereas others are barren.
Some of the earliest subdivisions of the Chalk (e.g. Barrois, 1876), used the abundance of particular groups of fossils, such as the echinoids Holaster (Cenomanian), and Micraster (Coniacian, Santonian), the crinoid Marsupites (Santonian) and belemnites (Campanian–Maastrichtian). These broad divisions are still useful concepts as they emphasize the conspicuous faunal assemblages present at different times, perhaps reflecting major ecological changes in the Upper Cretaceous oceans.
Key biostratigraphical indices
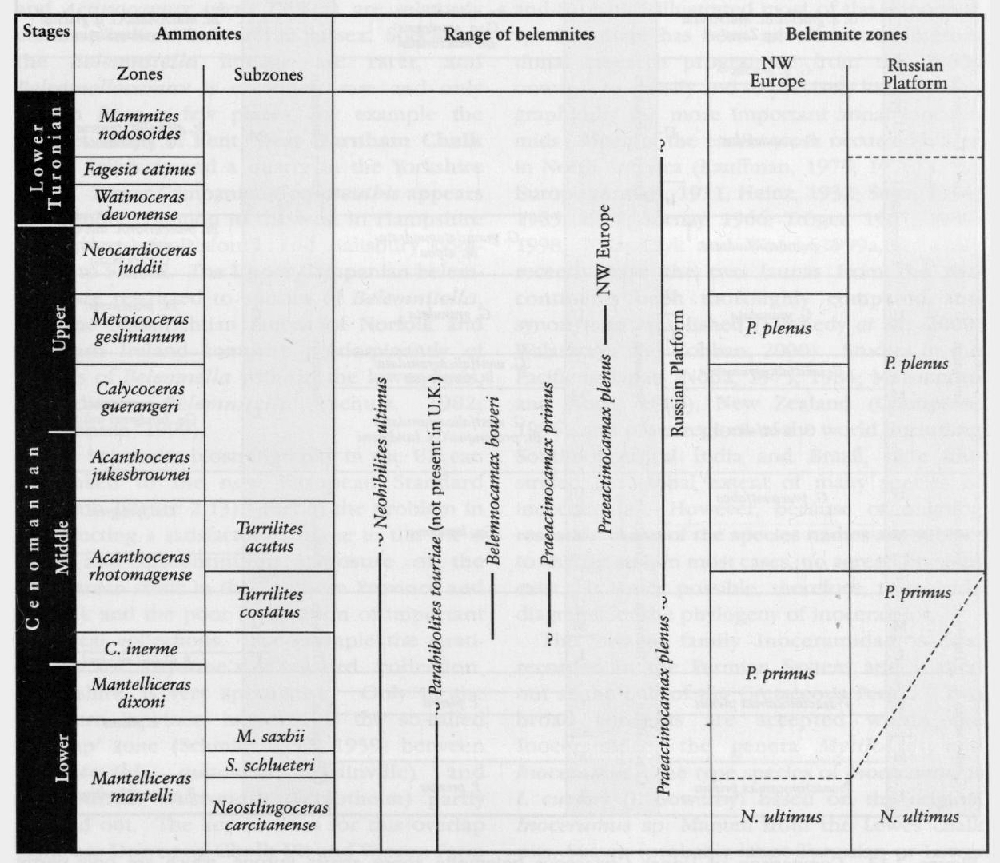
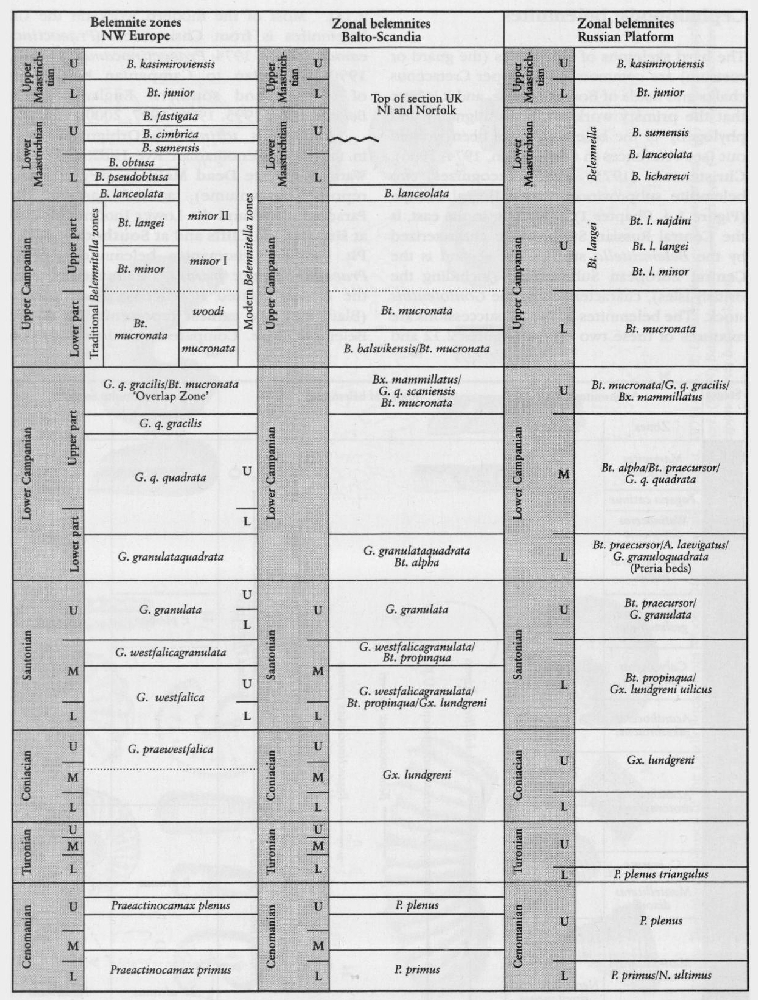
In the Upper Cretaceous Series, as in other parts of the Mesozoic succession, cephalopods, particularly the ammonites, are generally considered to be the most important international stratigraphical index fossils. This is because the original stage concepts are largely based on ammonites (see Appendix, this volume), and because of the widespread occurrence of ammonites in marine sediments. Ammonites are relatively common in the more marly chalks of the Cenomanian Stage and the condensed, shallow-water shelf areas of south-west England and the Anglo-Brabant Massif. In the purer chalks of the Turonian to Maastrichtian strata, ammonites are less common. In place of ammonites, the inoceramid bivalves are common throughout the Chalk and the value of these fossils for stratigraphy has become internationally recognized in the last two decades. Other cephalopods, such as the belemnites, are common at certain horizons representing incursions from Boreal northern Europe. Belemnites are a feature of the Chalk from the base of the Campanian Stage into the Maastrichtian Stage.
For local correlation, particularly in the Boreal Upper Cretaceous succession of Europe (
Cephalopods: ammonites
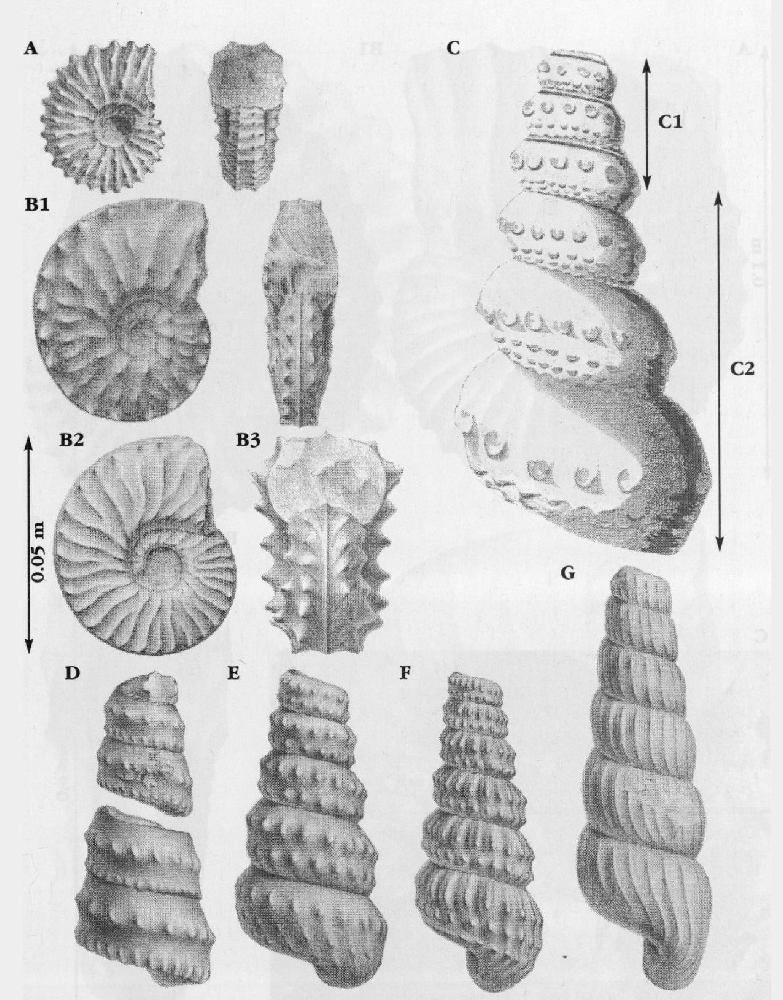
Sharpe (1853–1857) monographed the British Upper Cretaceous cephalopods (see examples in
The essentially Tethyan Vascoceratidae are less common in the English Chalk, but one species, Fagesia catinus (Mantell) is the index species of one of the Lower Turonian zones. The Collignoniceratidae include Collignoniceras woollgari (Mantel), and Subprionocyclus neptuni (Geinitz), the zonal index fossils of the Middle and the greater part of the Upper Turonian strata respectively.
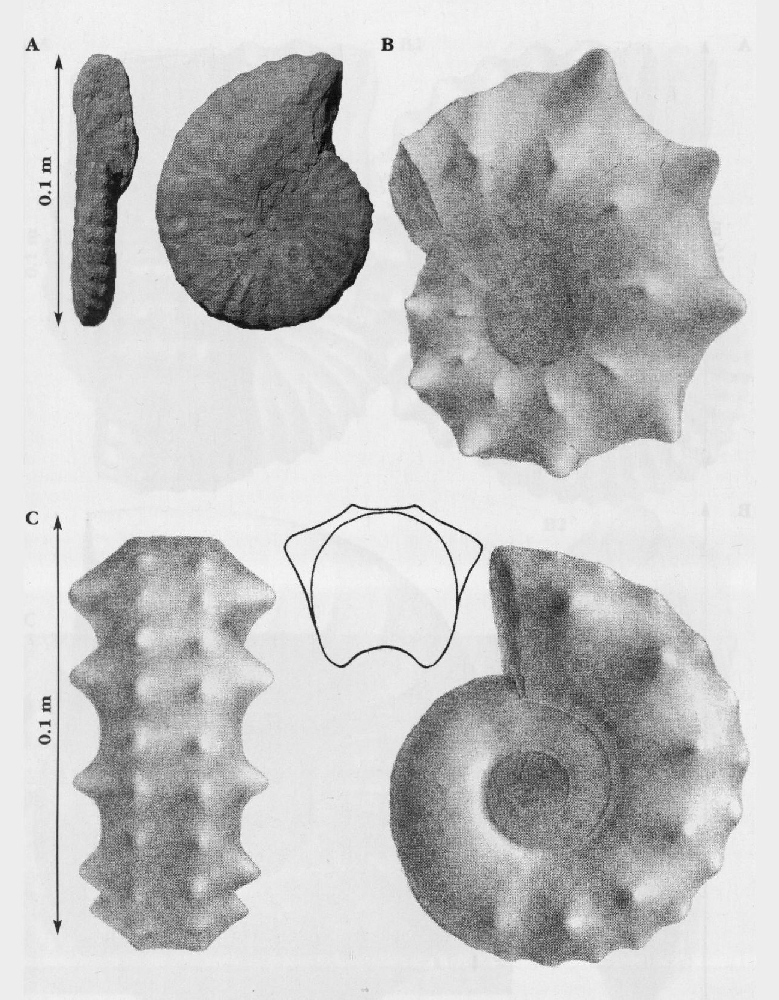
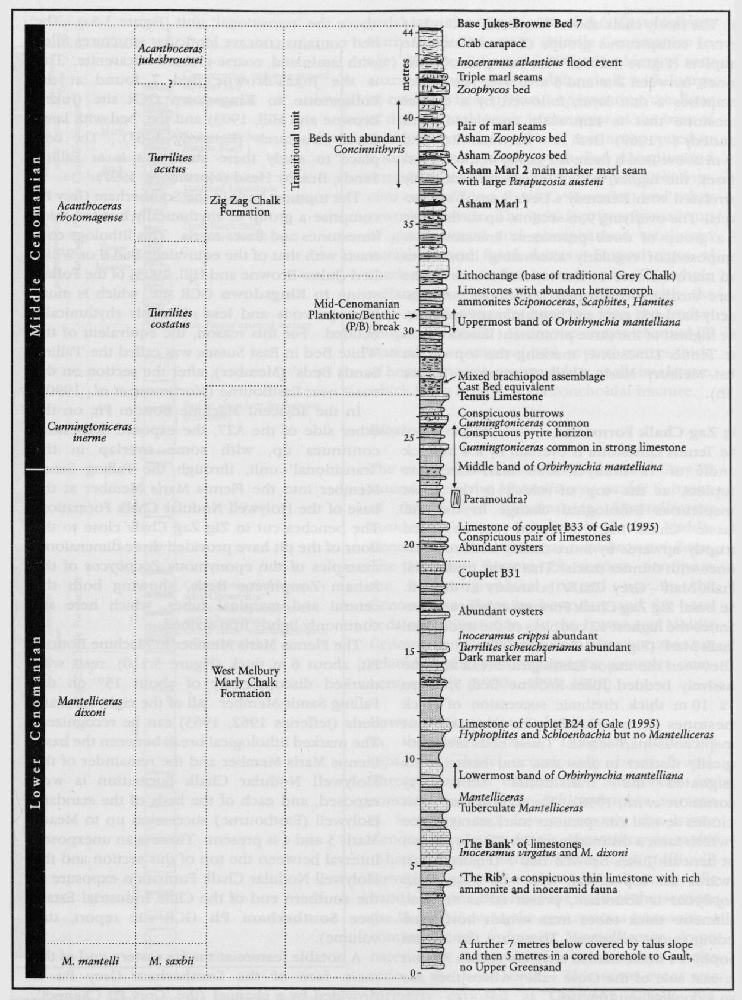
In addition to the main zonal and subzonal index fossils, the Cenomanian succession contains a spectacular variety of ammonites. The so-called heteromorph or uncoiled ammonites, including the genera Neostlingoceras, Algerites, Hypoturrilites, Mariella, Scaphites, Idiohamites, and Sciponoceras, are abundant at some levels, particularly as well-preserved, three-dimensional fossils in the hard limestones around the mid-Cenomanian (P/B) break
Very large desmoceratid ammonites (Parapuzosia (Austiniceras) austeni (Sharpe);
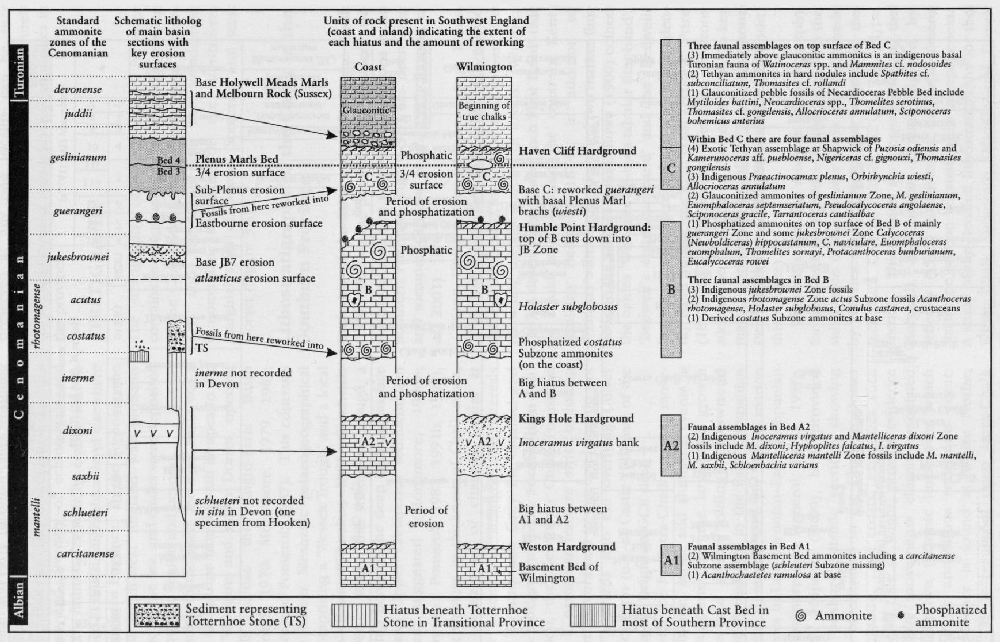
In much of Europe and particularly in the UK, the highest Cenomanian and lowest Turonian succession is highly condensed compared with areas such as North America. In this interval there are several ammonite zones represented in the UK by thin, condensed or reworked sediments, making recognition of the zones exceedingly difficult. It is only recently that the more expanded sections such as Eastbourne and Beachy Head have yielded ammonites of the Neocardioceras juddii, Watinoceras devonense and Fagesia catinus zones. In south-west England these ammonites are better preserved, but in such condensed successions that the relationship with more expanded sections has been difficult to establish (
Where well-preserved ammonites are found in otherwise ammonite-poor parts of the succession, even as single horizons, then the UK succession can be placed more confidently in the international zonal scheme. Two such horizons are the aragonitic fossils from the Upper Turonian Chalk Rock at Kensworth Chalk Pit Subprionocyclus neptuni Zone (Woods, 1896; Wright, 1979: Kaplan et al., 1987; see
Cephalopods: belemnites
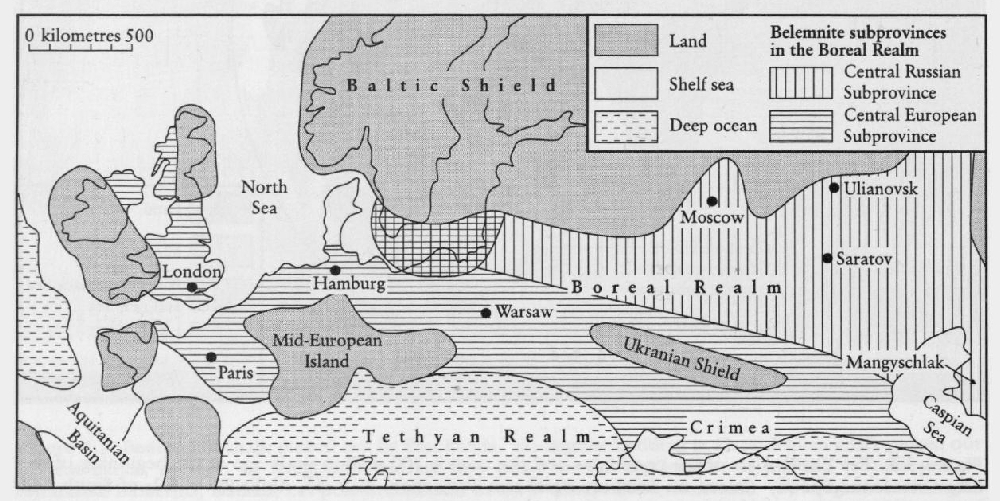
The hard skeletons of belemnites (the guard or rostrum) are common in the Upper Cretaceous chalks and marls of Boreal Europe, and it is here that the primary work on the stratigraphy and phylogeny of the belemnites has been worked out (see references in Christensen, 1974, 1986). Christensen (1975, 1986) recognizes two belemnite subprovinces within Boreal Europe (
Neohibolites ultimus (d'Orbigny) occurs in the basal Cenomanian Rye Hill Sands near Warminster (see Dead Maid Quarry GCR site report, this volume), at the base of the
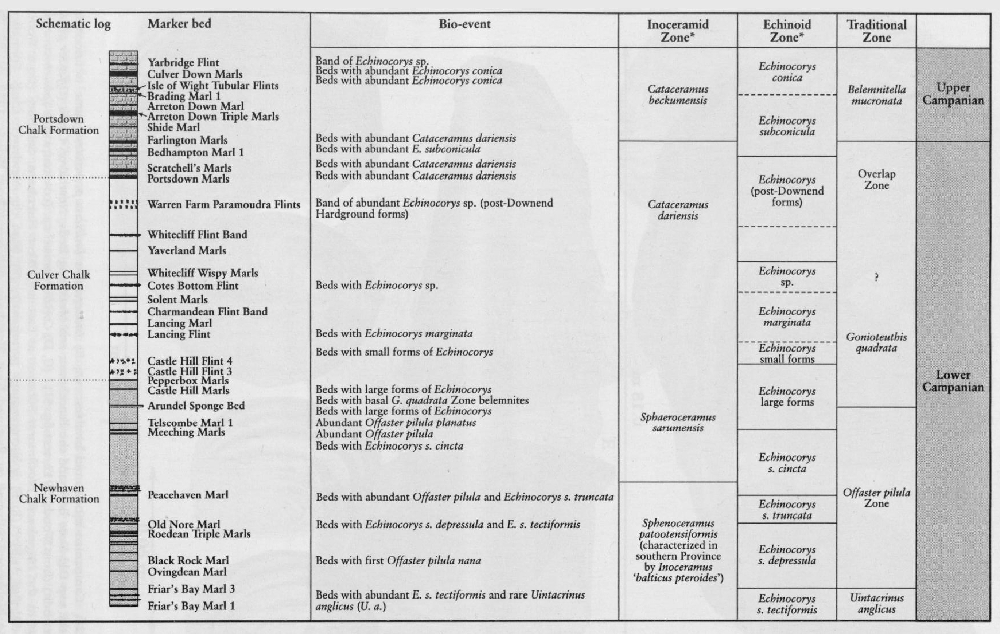
In the Coniacian to Lower Campanian chalksof England, species of the Gonioteuthis lineage and Actinocamax verus (Miller) are relatively common in Kent, but rarer in Sussex. Species of the Belemnitella lineage are rarer, and Belemnellocamax is extremely rare and only known from a few places, for example the Thanet Coast of Kent, West Harnham Chalk Pit in Salisbury and a quarry in the Yorkshire Wolds. Lower Campanian Gonioteuthis appears to be more common to the west in Hampshire (Portsdown, Mottisfont), and Salisbury compared to Sussex. The Upper Campanian belemnites are restricted to species of Belemnitella, and the Maastrichtian faunas of Norfolk and Northern Ireland comprise predominantly of species of Belemnella with, in the lower beds, subordinate Belemnitella (Schulz, 1982; Christensen, 1999).
The belemnite biostratigraphy in the UK can be linked to the new European Standard Zonation
Bivalves: inoceramids
Perhaps the most consistently abundant fossils in the Upper Cretaceous Chalk are the inoceramid bivalves. Even the hinges alone are frequently sufficiently distinctive to have biostratigraphical value. These bivalves exhibit a great variety of forms, which are generally easily identifiable, and have restricted ranges, providing ideal zonal material. Following the monograph of Woods (1911–1912, Volume 2, Parts 7 and 8), which illustrated most of the important species, there has been an enormous international research programme from the 1960s onwards to classify and to precisely locate stratigraphically the more important zonal inoceramids. Much of the earlier work occurred either in North America (Kauffman, 1975, 1976) or in Europe (Andert, 1911, Heinz, 1932; Seitz, 1934, 1965, 1967; Sornay, 1966; Trtiger, 1967, 1989, 1998; Walaszczyk and Wood, 1999a,c). Only recently have the two faunas from the two continents been thoroughly compared and synonymies established (Kennedy et al., 2000; Walaszczyk and Cobban, 2000). Studies in the Pacific in Japan (Noda, 1975, 1984; Matsumoto and Noda, 1986), New Zealand (Crampton, 1996), and other regions of the world, including Southern Africa, India and Brazil, have illustrated the global extent of many species of inoceramids. However, because of ongoing research, many of the species names are subject to change and, in most cases, no agreed lineages exist. It is not possible, therefore, to provide diagrams for the phylogeny of inoceramids.
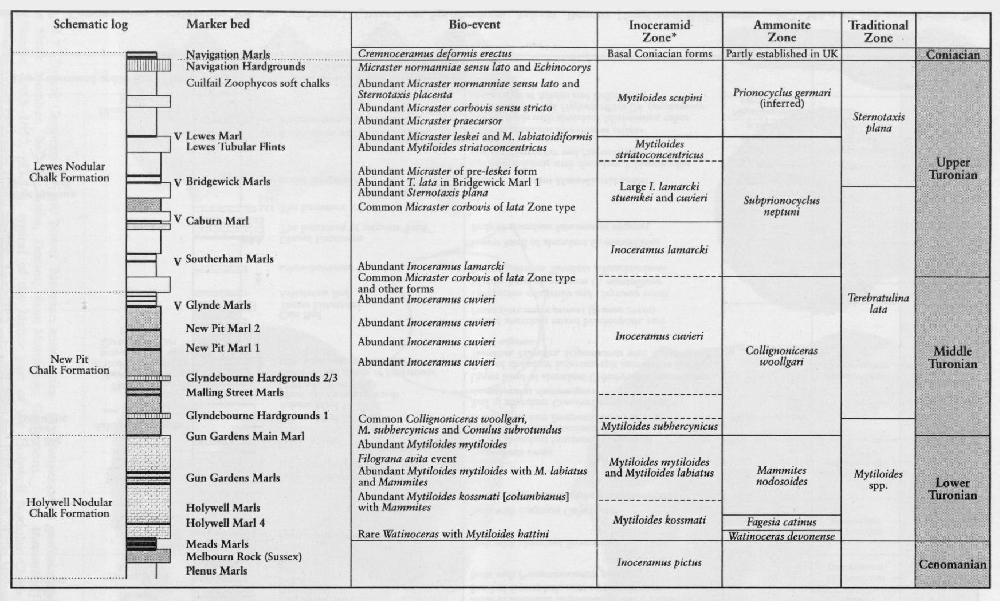
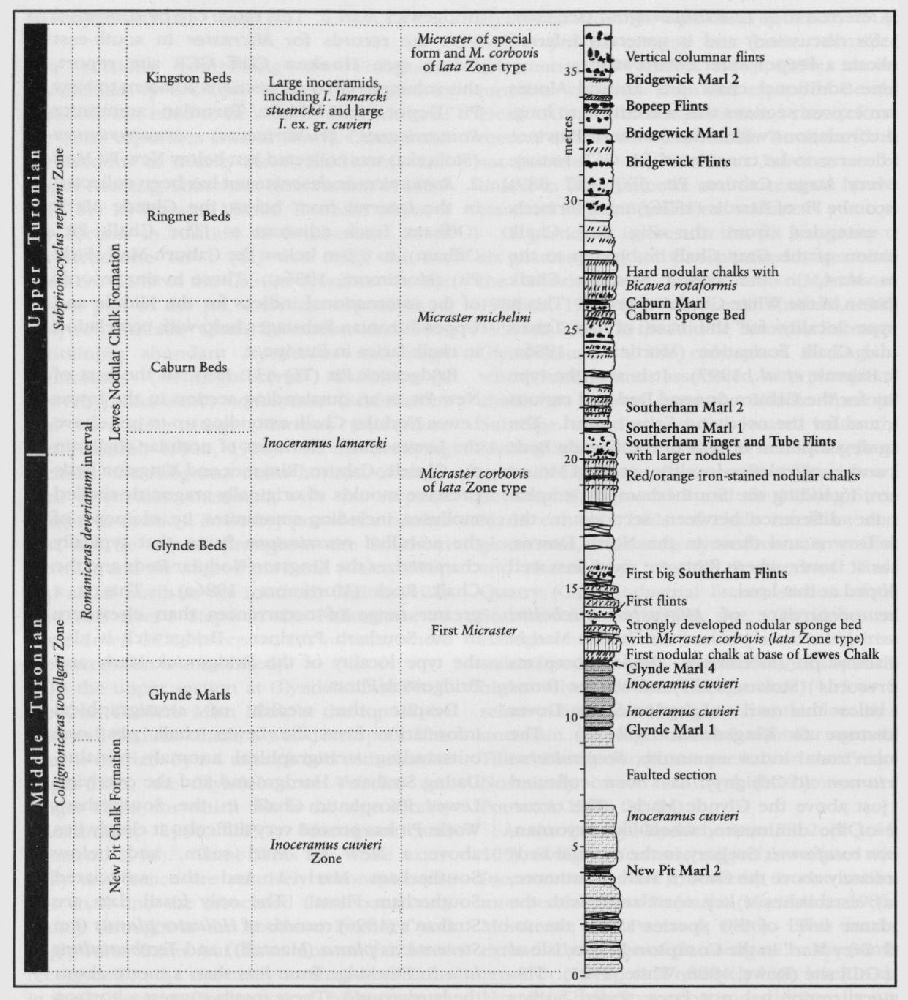
The bivalve family Inoceramidae is first recorded in the Permian System and it died out at the end of the Cretaceous Period. Two broad concepts are accepted within the Inoceramidae, the genera Mytiloides and Inoceramus. The type species of Inoceramus is I. cuvieri (J. Sowerby) based on the original Inoceramus sp. Mantell from the Lewes chalk pits, Sussex (probably latest Turonian or Lower Coniacian in age from the Southerham Pit). Mytiloides is essentially a Turonian genus whereas Inoceramus ranges throughout the Upper Cretaceous succession. Other genera include Cremnoceramus and Volviceramus (Coniacian), Cladoceramus (Santonian), Sphenoceramus (Santonian–Campanian) and Cataceramus (Campanian).
Inoceramids tend to occur in great abundance at particular horizons. The Cenomanian species Inoceramus crippsi (Mantell)
The most troublesome horizon for stratigraphers and palaeontologists has been that yielding the Lower Turonian Mytiloides assemblages
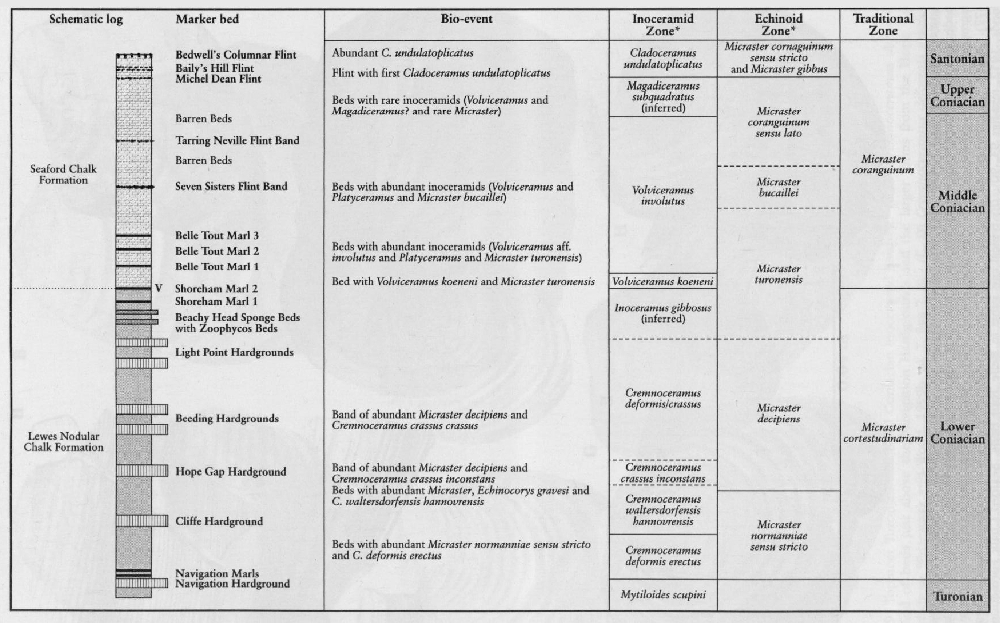
The Second Inoceramid Workshop (held in Freiburg in 1996; Walaszczyk and Wood, 1999a) concentrated on the Upper Turonian and Lower and Middle Coniacian assemblages
In the Middle Coniacian to Upper Santonian interval, the occurrence and ranges of inoceramid species are comparatively well known
Other bivalves
The Upper Cretaceous Chalk is rich in bivalves other than inoceramids. Oysters are common at many horizons and are used for correlation in the Cenomanian succession (Pycnodonte events, see
The book Fossils of the Chalk (Owen and Smith, 1987) contains excellent illustrations of many of the more important fossil bivalves.
Brachiopods
Brachiopods are surprisingly abundant throughout the Upper Cretaceous Chalk. There are some horizons where brachiopods are the key indices for correlation and provide an additional environmental indicator of conditions on the chalk seabed. Both inarticulate and articulate brachiopod classes are present, the Inarticulata being represented by the long-ranging genera Ancistrocrania and Isocrania. By far the most abundant brachiopods in the Chalk are the Articulata with the genera Orbirhynchia, Cretirhynchia, Gibbithyris, Concinnithyris and Terebratulina being the most widely used for stratigraphy. Following Davidson's monographs of British Brachiopoda (1852–1854, 1874), the Chalk Terebratulidae were described by Sahni (1929) and the Rhychonellidae were described by Pettitt (1949, 1954). These remain major references for the Upper Cretaceous brachiopods. Owen (1962, 1968, 1970, 1977, 1988) revised many Upper Cretaceous genera. Owen also illustrated all the major species in his book Fossils of the Chalk (1987).
There are two key aspects to the Chalk brachiopods. The first relates to stratigraphical shifts in diversity and the second to 'pioneering' species that are the first indicators of change in the environment. There are also major shifts in size at some horizons.
In the Cenomanian many chalks, horizons with diverse brachiopod assemblages, such as the
A marked change in brachiopod size occurs across the
At many horizons it is the brachiopods that are key guide fossils. These include the three Orbirhynchia mantelliana events in the Cenomanian Stage, the O. multicostata (Pettitt) event in the Plenus Marls Bed 1, and O. dispansa (Pettitt) in the Bridgewick Marls and the Turonian Stage, and Magas chitoniformis (Schlotheim) in the higher part of the Upper Campanian and Lower Maastrichtian substages.
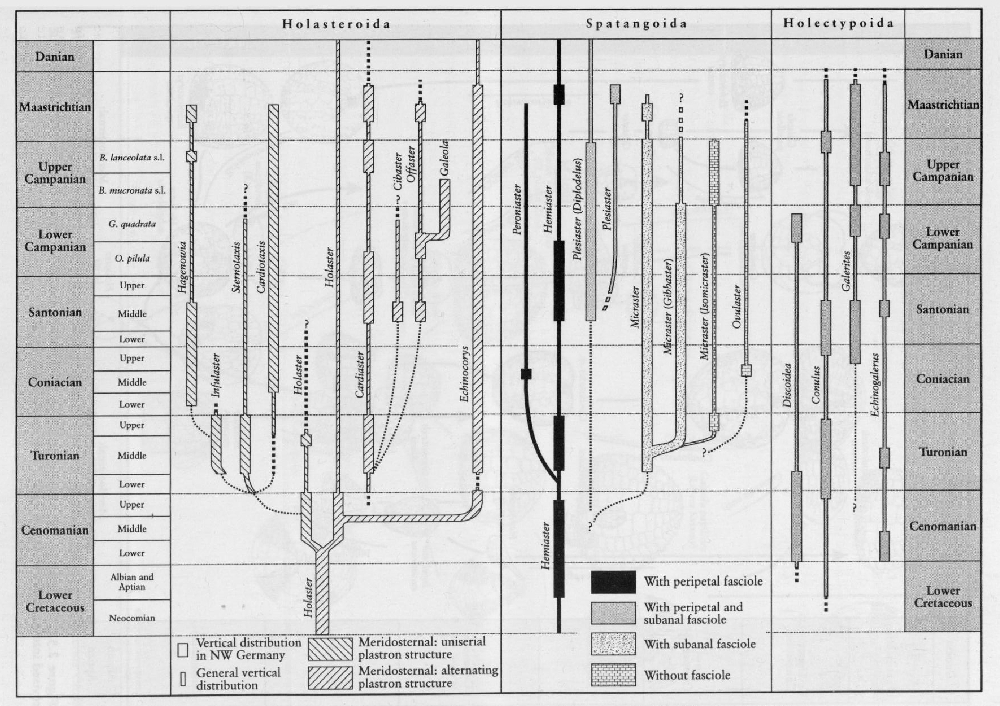
Sea urchins: the echinoids
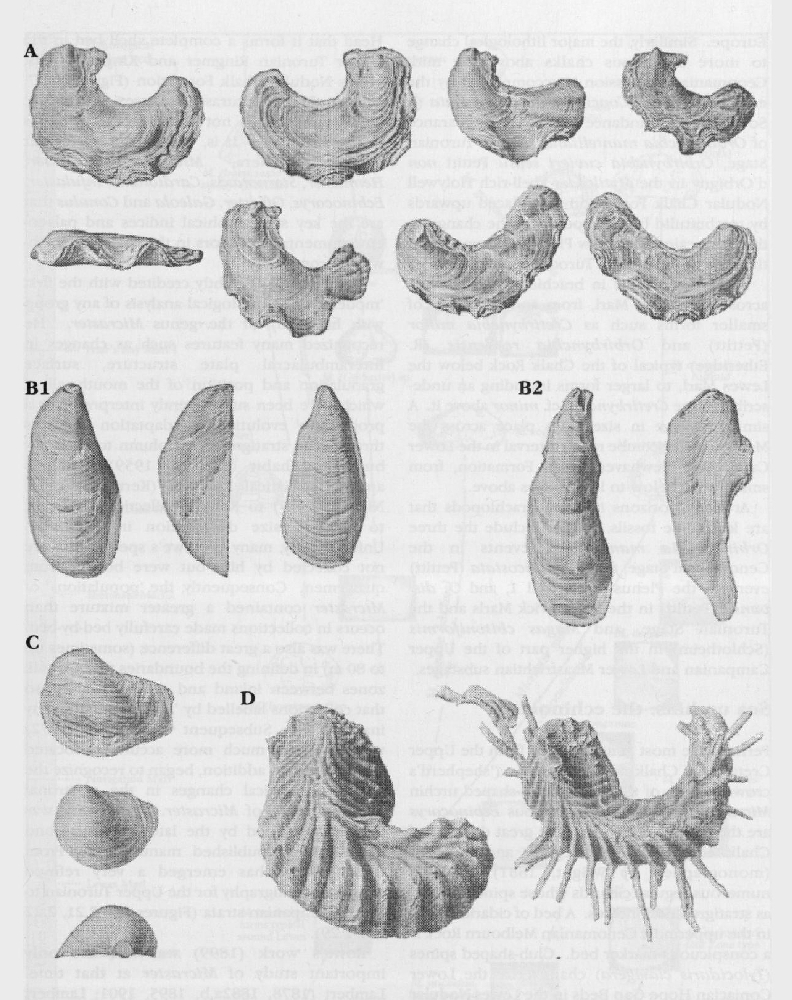
Perhaps the most famous fossils from the Upper Cretaceous Chalk are sea urchins ('shepherd's crowns') and, of these, the heart-shaped urchin Micraster and the less glamorous Echinocorys are the most well known. The great diversity of Chalk echinoids, both regular and irregular (monographed by Wright, 1881), includes numerous regular cidarids whose spines also act as stratigraphical indices. A bed of cidarid spines in the uppermost Cenomanian Melbourn Rock is a conspicuous marker bed. Club-shaped spines Tylocidaris clavigera) characterize the Lower Coniacian Hope Gap Beds in the
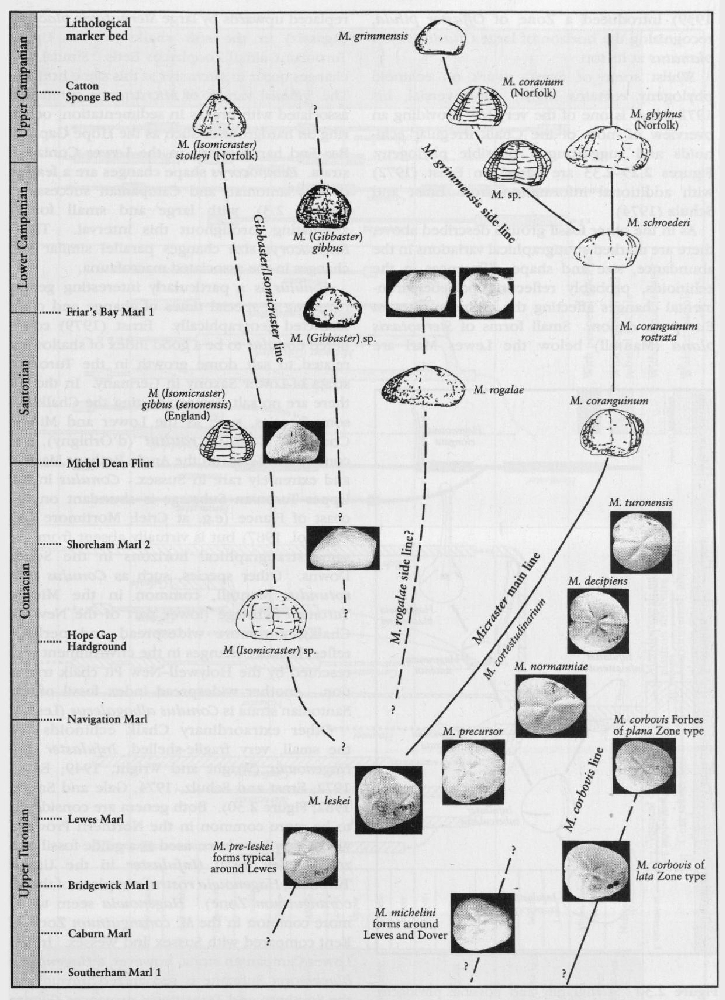
Rowe (1899) is rightly credited with the first 'modern' palaeontological analysis of any group with his study of the genus Micraster. He recognized many features such as changes in interambulacral plate structure, surface granulation and position of the mouth, all of which have been subsequently interpreted as a progressive evolutionary adaptation upwards through the stratigraphical column to a deeper burrowing habit (Nichols, 1959). Others applied statistical analyses (Kermack, 1954; Nichols, 1959) to Rowe's collection and tried to show a size distribution in Micraster. Unfortunately, many of Rowe's specimens were not collected by him but were bought from quarrymen. Consequently, the 'populations' of Micraster contained a greater mixture than occurs in collections made carefully bed-by-bed. There was also a great difference (sometimes up to 80 m) in defining the boundaries to the Chalk zones between inland and coastal sections so that collections labelled by 'Zone' contain many inaccuracies. Subsequent work (Ernst, 1972) was based on much more accurately located samples and, in addition, began to recognize the key stratigraphical changes in the plastronal plate structure of Micraster. This work was further developed by the late Dr Drummond (1983, and unpublished manuscript). From these studies has emerged a very refined Micraster stratigraphy for the Upper Turonian to Lower Campanian strata
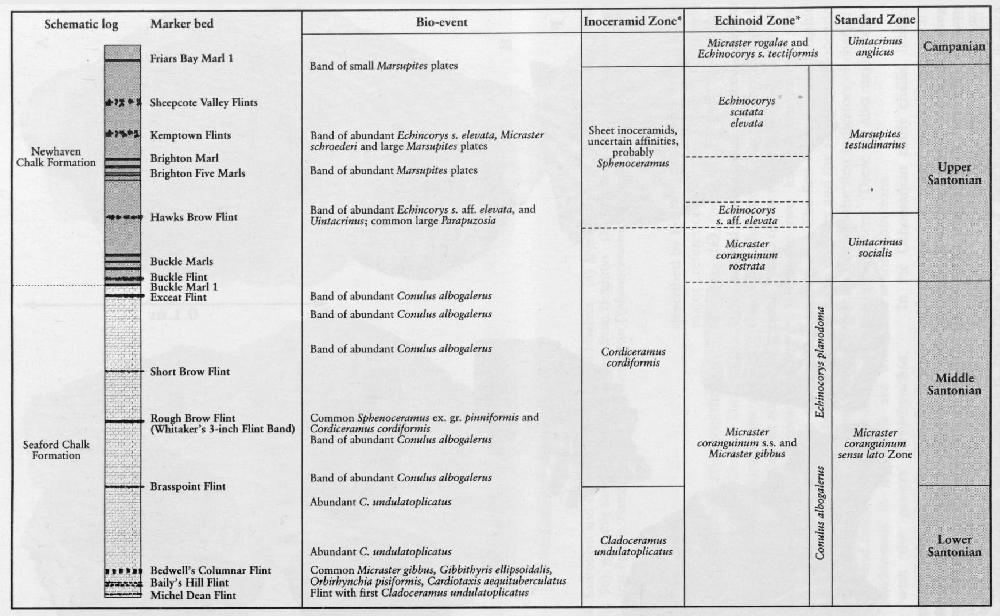
Rowe's work (1899) was not the only important study of Micraster at that time. Lambert (1878, 1882a,b, 1895, 1901; Lambert and Thiery, 1924) described the Micraster from the eastern Paris Basin and other echinoids in the Chalk including Echinocorys. Brydone (1912, 1914, 1915, 1930) and Gaster (1924) also recognized the stratigraphical importance of shape changes in Echinocorys. Brydone (1912, 1939) introduced a Zone of Offaster pilula, recognizing the horizon of large Offaster pilula planatus at its top.
Whilst some of Ernst's work on echinoid phylogeny remains highly controversial, his 1972 paper is one of the very few providing an overview of most of the Chalk irregular echinoids and suggesting a possible phylogeny.
As in the other fossil groups described above, there are marked stratigraphical variations in the abundance, size and shape differences in the echinoids, probably reflecting palaeoenvironmental changes affecting the entire north-west European region. Small forms of Sternotaxis plana (Mantell) below the
Conulus is a particularly interesting genus, recurring at special times of change and often restricted geographically. Ernst (1979) considered Conulus to be a good index of shallowing related to salt dome growth in the Turonian strata of Lower Saxony in Germany. In the UK there are no salt domes affecting the Chalk but some species, such as the Lower and Middle Coniacian Conulus raulini (d'Orbigny), are common in Kent (on the Anglo-Brabant Massif), and extremely rare in Sussex. Conulus in the Upper Turonian Substage is abundant on the coast of France (e.g. at Criel; Mortimore and Pomerol, 1987) but is virtually absent from the same stratigraphical horizons in the South Downs. Other species, such as Conulus subrotundus Mantell, common in the Middle Turonian Substage (lower part of the New Pit Chalk), are more widespread and perhaps reflect broader changes in the environment represented by the Holywell–New Pit chalk transition. Another widespread index fossil of the Santonian strata is Conulus albogalerus (Leske).
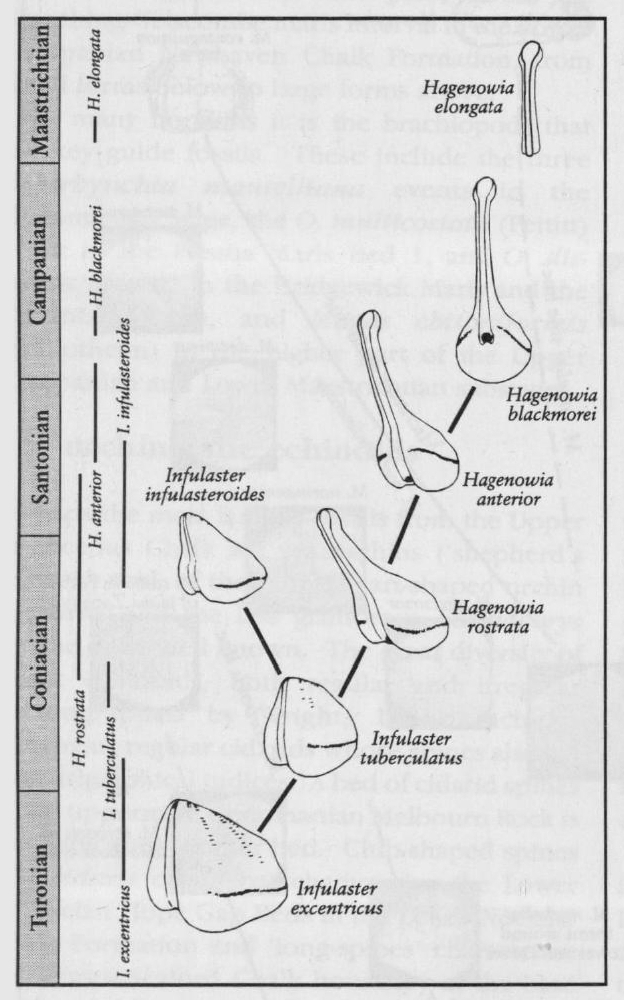
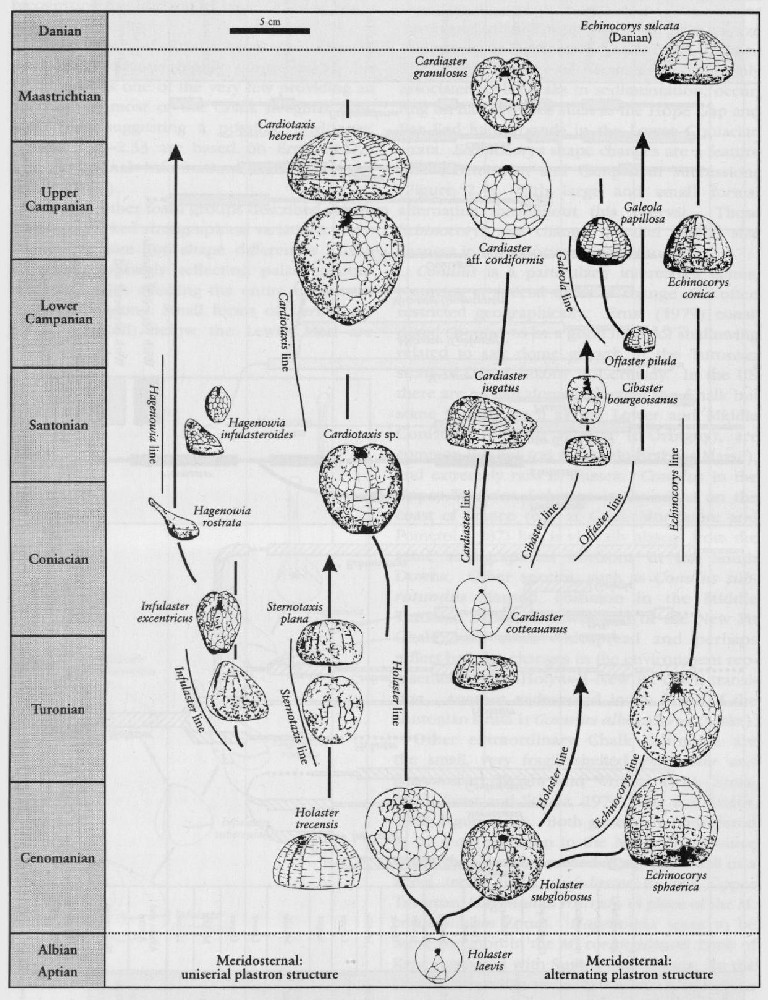
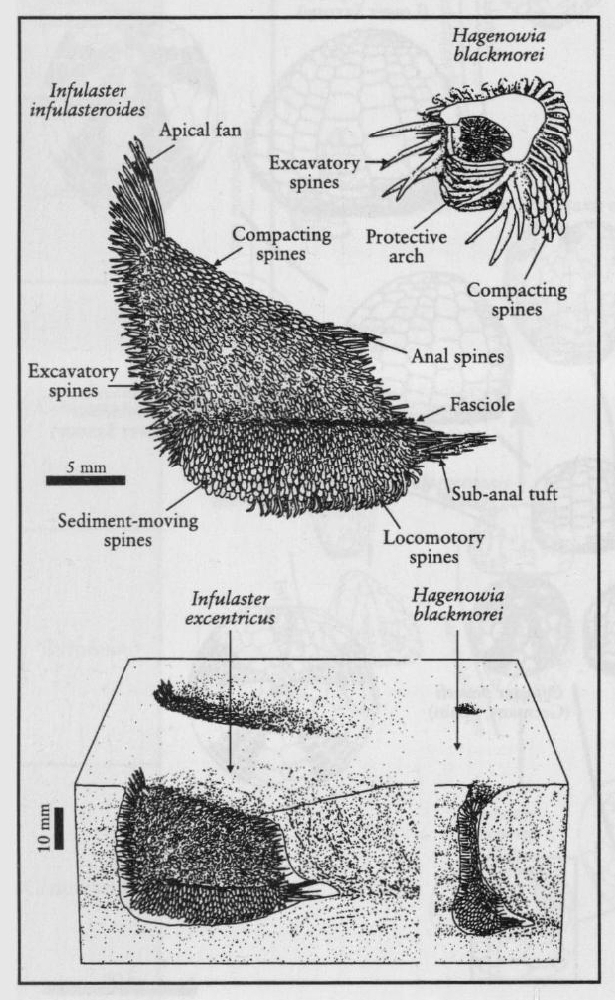
Other extraordinary Chalk echinoids are the small, very fragile-shelled, Infulaster and Hagenowia (Wright and Wright, 1949; Ernst, 1972; Ernst and Schulz, 1974; Gale and Smith, 1982;
Sea lilies and feather stars: the crinoids
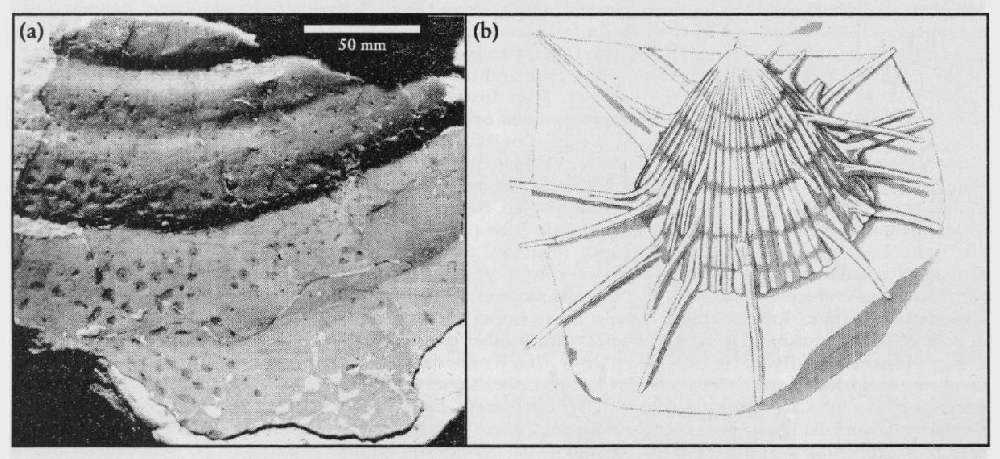
The marvellous drawings of Upper Cretaceous Chalk crinoids in books such as Dixon's Geology of Sussex (1850) indicate both the variety and the type of preservation once found in hand-dug chalk pits. The two main types of crinoid, sea lilies (sessile, with stems) and feather stars (pelagic, stemless) are both present in the Chalk. Crinoids are very common at some horizons and are crucial zonal index fossils at other horizons. The
The worldwide occurrence of the stemless but benthic crinoids Uintacrinus socialis Grimmell, U. anglicus Rasmussen and Marsupites testudinarius Schlotheim makes them key guide fossils. It is unusual to have the last occurrence of a species as a stage boundary index, but such is the consistent and abundant occurrence of Marsupites at its extinction point that there is nothing else to better it. Uintacrinus socialis is most abundant in two bands, one at the base and one at the top of its range. Marsupites is relatively abundant throughout its range but has two bands of maximum abundance. There are also two stratigraphically distinct morphotypes of Marsupites. The first morphotype, in the basal part of the Marsupites testudinarius Zone, has calyx plates with a simple fold in the centre of each edge, whereas the second morphotype has complex plates with complex folds providing crinkled edges. There is also a stratigraphically useful size change in Marsupites, which tends to be smaller at the base and top of its range.
Trace fossils
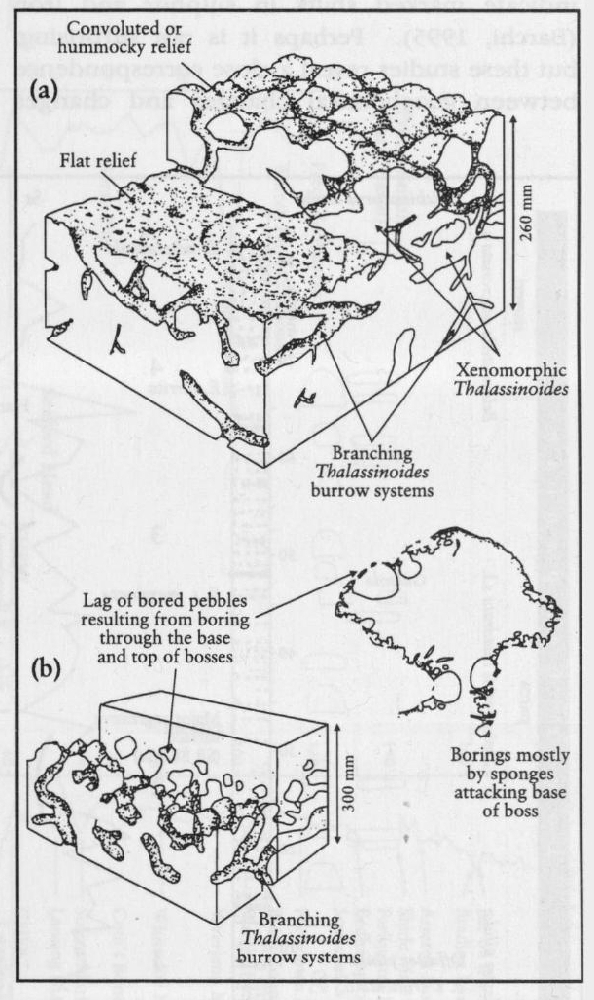
Many animals are very rarely preserved as fossils in the Chalk. The evidence from the abundance of trace fossils is that the seabed was inhabited by a wide range of animals, some of unknown affinities (Bromley, 1990). The branching trace fossil Thalassinoides is ubiquitous in the Chalk, and Bromley (1967, 1975a;
Other trace fossils such as Zoophycos, Chondrites and Bathichnus paramoudrae
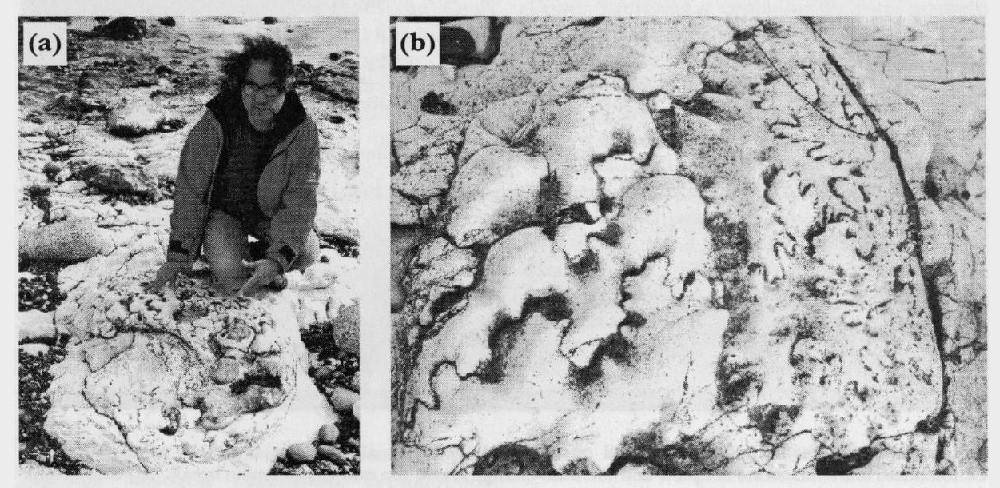
Just as body fossils have levels of abundance used for stratigraphical correlation, so the trace fossils have conspicuous horizons of abundance. These include the abundant Zoophycos levels, called 'Tiger' chalk in the Crimea and Banderkreide in Germany. Similarly, Paramoudra is stratigraphically restricted (Mortimore and Pomerol, 1991b).
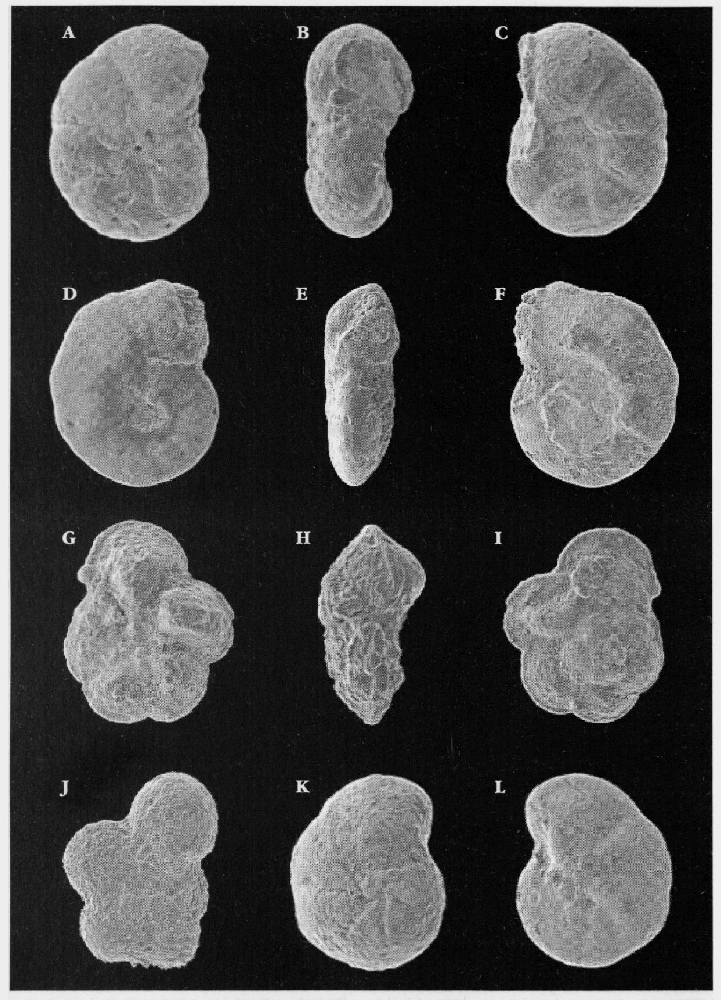
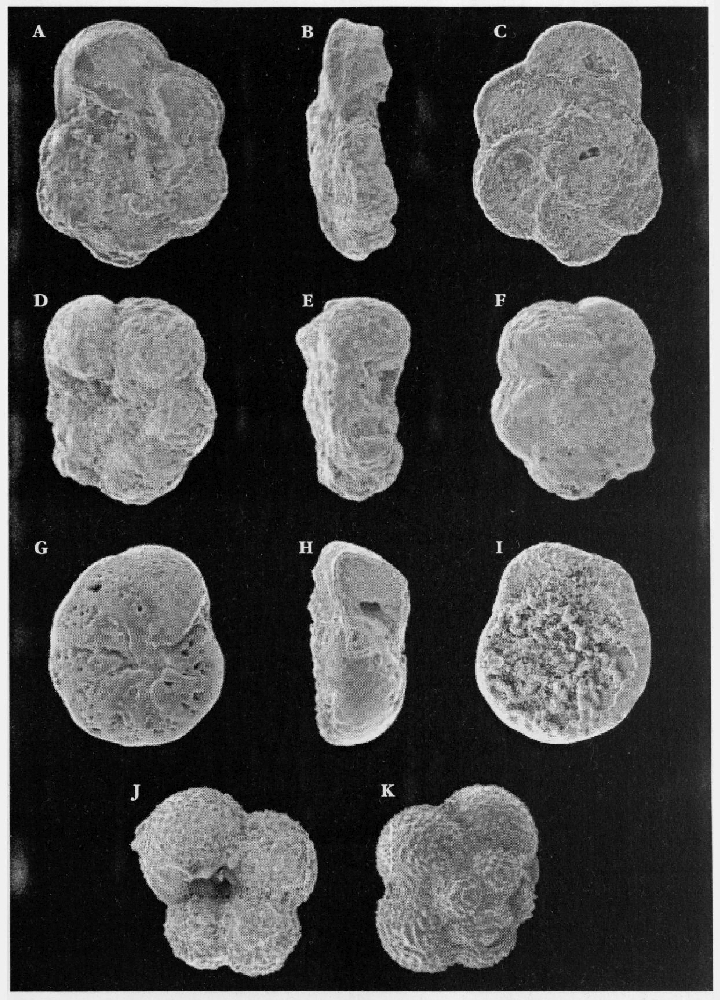
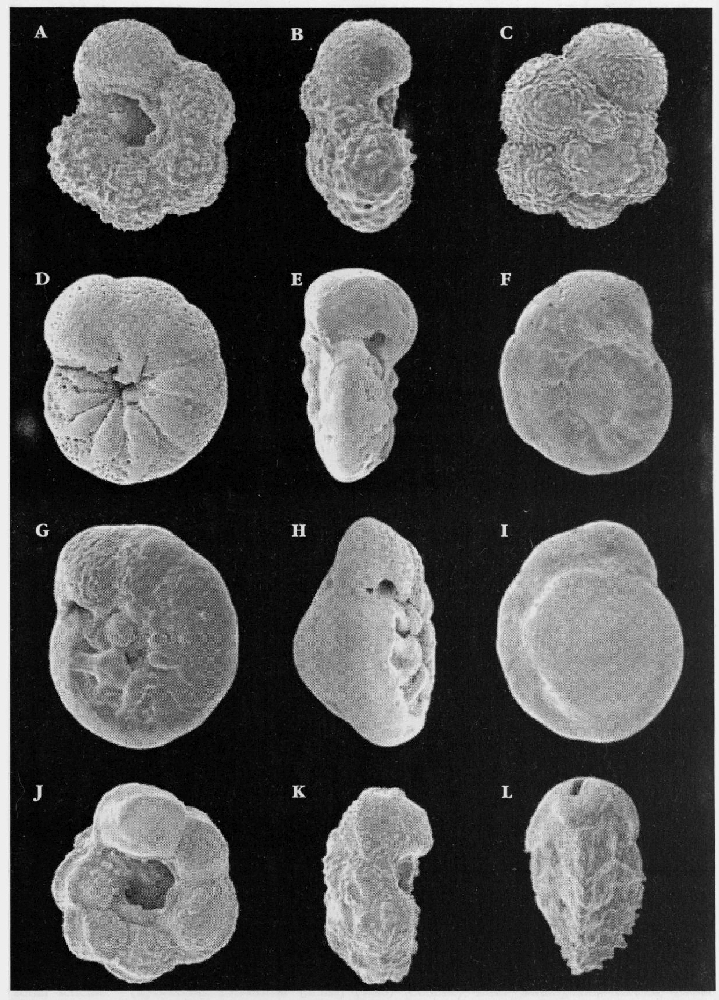
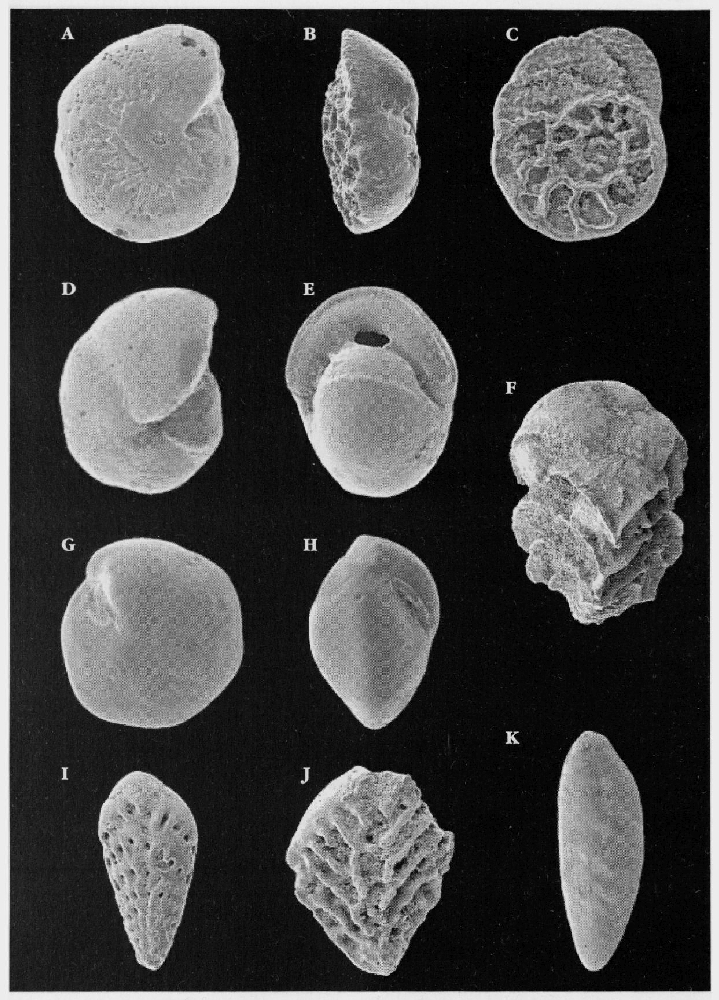
Microfossils
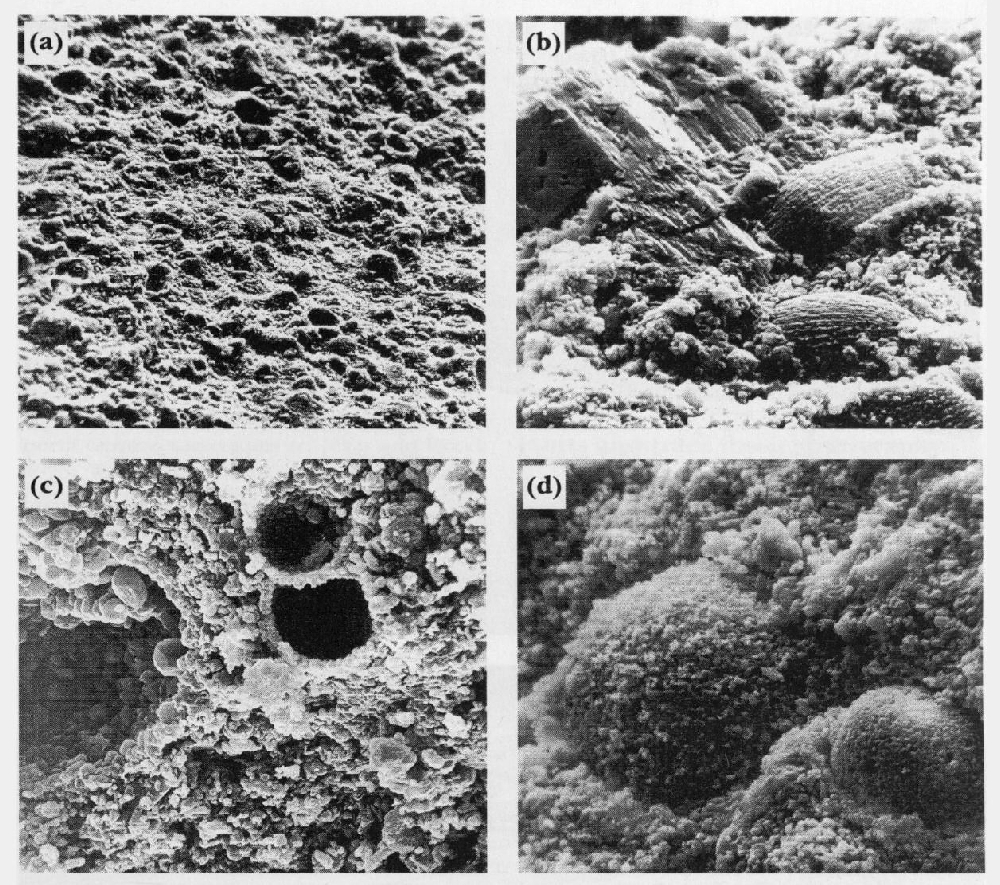
Upper Cretaceous soft chalks are full of well-preserved microfossils
The ecology of the Chalk seas
There are environmental effects on fossil distribution and preservation before the chalk sediment even forms a substrate on the seabed. Chalk is made primarily from pelagic nannofossils (especially coccoliths;
The evidence from both trace fossils (Bromley, 1975a) and from the type of granulation on the tests of echinoids such as Hagenowia (Gale and Smith, 1982), indicate that the initial 'chalky-soup' became a partly consolidated soft- to firm-ground relatively rapidly. Only a millimetre to centimetre thin layer was probably available for re-suspension at the sediment–water interface. This was the environment for soft-sediment colonizers. Some burrowed into and through the sediment (e.g. Hagenowia,
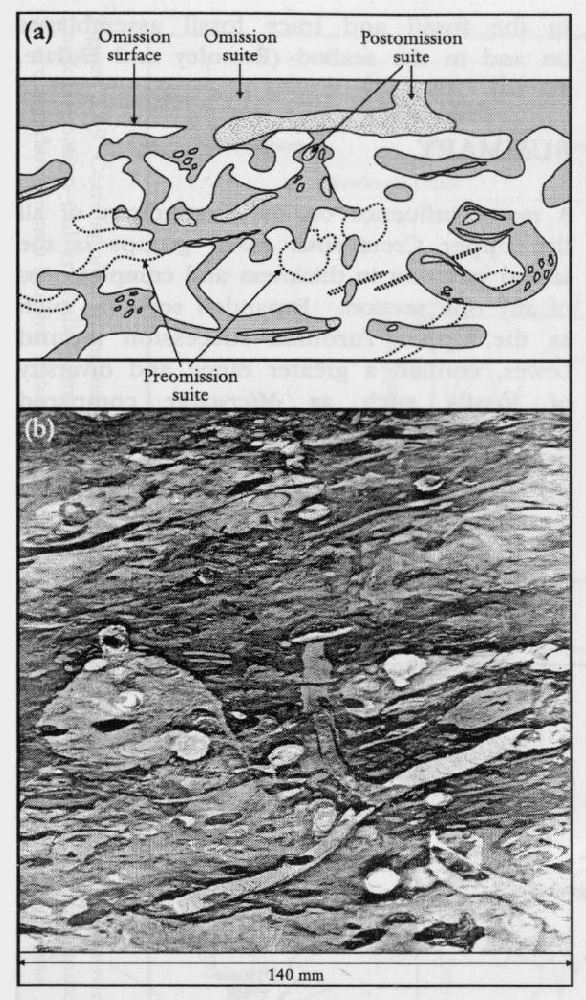
Some surfaces on the seabed progressed from soft- to firm- to hardgrounds
Westermann (1990) identified the ecological niches occupied by Mesozoic (including Upper Cretaceous) ammonites (Wiedmann, 1996). Heteromorph ammonites such as the turrilitids and scaphitids might represent shallow water grazers on algae or seaweed, an idea also proposed by others such as Drummond (1967). The large parapuzosid ammonites may have been adapted to a greater range of depths.
In contrast to the deeper-water chalks, a much greater diversity of fossils is found on shallower water shelf areas (e.g. Wilmington Quarry) of south-west England during the Cenomanian Age, where calcareous sands were the substrate.
The inoceramids in particular are thought to have been especially adapted, partly by chemosymbioses, to oxygen-poor conditions on the seabed including Mesozoic 'black-shale' events (Kauffman, 1988; Kauffman and Harries, 1992). The low-diversity Mytiloides labiatus sensu lato fauna in the Holywell Nodular Chalk is cited as one example of this adaptation and these special seabed conditions may be related to major sea-level fluctuations.
Integrated ecology; body fossils, trace fossils and isotopes
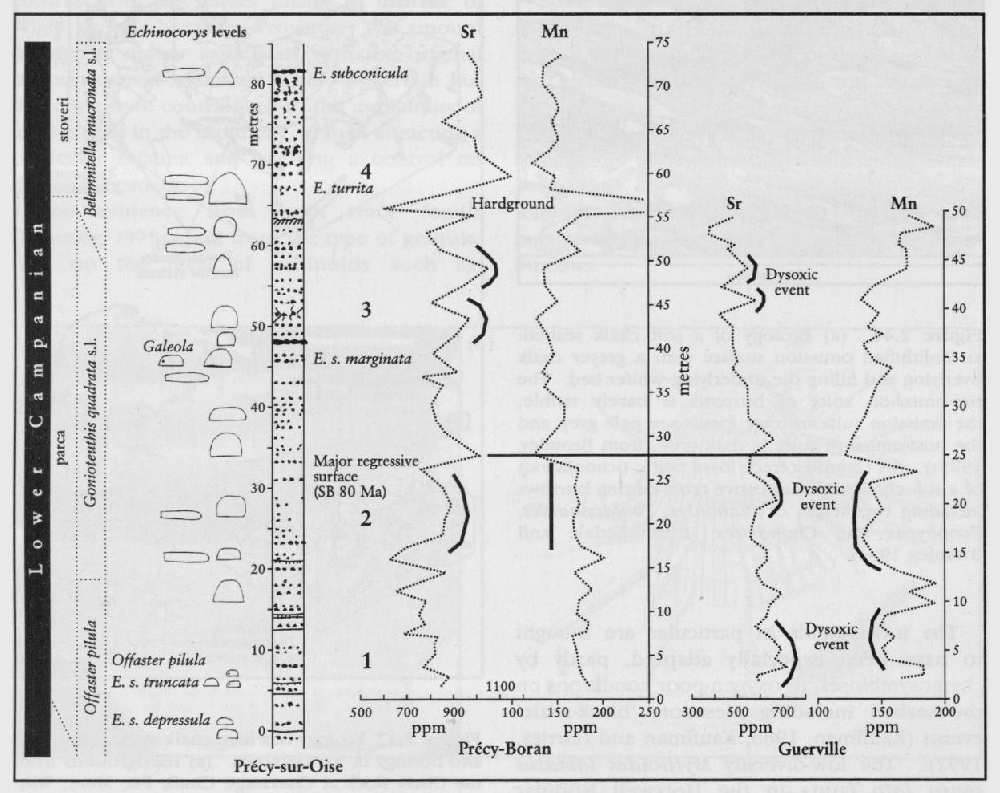
By combining the evidence from the stratigraphical and geographical distribution of body fossils and trace fossils a picture of the ecology of the Chalk seabed can be constructed. In addition, studies of the stable isotopes of oxygen and carbon have identified the changes in seawater temperature (Jenkyns et al., 1994) during the Upper Cretaceous Epoch, and the major shifts in burial of organic carbon. Other studies of the stratigraphical distribution of elements
Summary
A major influence on the stratigraphy of all the Upper Cretaceous fossil groups is the lateral variation in thickness and completeness of any one section. Expanded sections, such as the Upper Turonian succession around Lewes, contain a greater range and diversity of fossils such as Micraster compared with more condensed sections in Kent and Wessex. Major erosional channels have reduced the completeness of the stratigraphy at many localities. The Upper Santonian succession of the Anglo-Brabant Massif on the Isle of Thanet and in East Anglia has a different distribution of belemnites and brachiopods compared with the deeper basinal sections of Sussex These aspects of the Chalk have been only recently recognized so that 'zonal' labels for fossil collections in museums can be highly misleading. It is only with a greater understanding of chalk sedimentary processes and their tectonic and sea-level controls, that the study of the palaeontology and stratigraphy of the Upper Cretaceous Epoch can be accomplished more satisfactorily. Ever more detailed studies are required if the spatial and temporal distribution of Chalk fossils is to be fully understood. Perhaps in years to come it will be possible to show the migration rates of Upper Cretaceous animals within individual cycles, across individual basins. The GCR sites described in this volume will be major references for such studies.